1. Introduction
Allergies affect millions of people worldwide and are characterized by an excessive type 2 immune response to normally harmless substances, generally known as antigens or allergens, specifically [
1,
2]. Consequently, this response leads to the development of various allergic symptoms, including asthma, allergic rhinitis, and atopic dermatitis. In the most severe cases, it can result in anaphylaxis and possibly death. According to the World Allergy Organization, the prevalence of allergic diseases has been continuously increasing in the industrialized world [
3,
4]. In addition, according to the World Health Organization, the number of asthma patients is expected to increase to 400 million by 2025 [
4]. The process by which the immune system becomes sensitive to a particular allergen is called sensitization and is typically accompanied by the development of immunoglobulin E (IgE), a specific subclass of antibodies, against the allergen. Sensitization rates to one or more common allergens among schoolchildren are reported to be between 40% and 50% [
5]. Since antigen E was isolated from the pollen of common ragweed (
Ambrosia artemisiifolia) as the first antigen in 1962 [
6], a variety of environmental and food allergens have been identified, including 106 allergens that have recently (between January 2019 and March 2021) been accepted by the Allergen Nomenclature Sub-Committee (
http://allergen.org/committee.php, accessed on 5 July 2023) [
7].
Antihistamines are widely used for symptomatic treatment of many allergic diseases with variable efficacy. Despite the identification of increasing varieties of antigens, there is no fundamental treatment to overcome allergic symptoms except for allergen immunotherapy or desensitization, whereby long-term remission is expected, against a few food and environmental allergens, e.g., cedar pollen [
12]. Desensitization therapy is actively investigated because of its clinical potential; however, it harbors the intrinsic risk of inducing severe side effects such as anaphylaxis [
13]. Thus, desensitization shots are co-administered with antihistamines. A drug that promotes immune tolerance by targeting macrophages, for instance, could make allergen desensitization safer and even more effective. In 2003, omalizumab, an IgE-blocking antibody received approval from the U.S. Food and Drug Administration (FDA), but it is not approved for all allergic conditions, and it is expensive [
14]. Alternatively, blocking antibodies against specific allergens are being developed, but these will be even more expensive for patients that are allergic to multiple allergens. Therefore, an alternative remedy based on a new concept is desirable for this growing patient population.
Immune cells such as mast cells, basophils, dendritic cells, B cells, and specific T-cell subsets are well recognized as key players in allergic reactions. In contrast, to date, macrophages are not commonly associated with allergies. In the future, it would be key for the field to provide credible in vivo evidence that macrophages also play a role in modulating allergy first using mouse models but ultimately in human patients. However, several lines of evidence have recently revealed the crucial role of macrophages in developing and modulating these allergic responses [
15,
16,
17,
18,
19,
20]. For example, macrophages are the most abundant immune cells present in the lungs (approximately 70% of the immune cells) and play a crucial role in asthma caused by environmental-allergen-induced airway inflammation [
21,
22], suggesting that macrophages, together with other immune cells could play a role in immune responses. Therefore, the role of macrophages in allergic diseases and the mechanism underlying their functional regulation deserve further study.
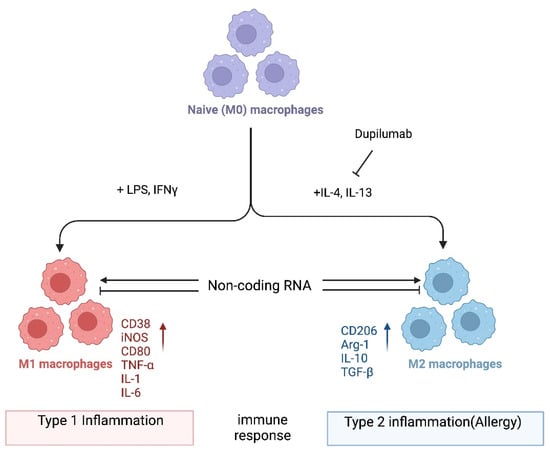
In one popular paradigm, macrophages can be divided into two major subclasses, i.e., M1 and M2, based on the inflammatory responses that they mediate, and the process by which macrophages differentiate in response to challenge is called macrophage polarization. Macrophage polarization is determined by the microenvironment (
Figure 1). However, the mechanisms underlying in vivo macrophage polarization are complicated and remain largely unclarified, but various intracellular molecules, including signaling molecules and enzymes, and receptors have been shown to regulate macrophage polarization [
23,
24,
25,
26,
27,
28,
29,
30,
31,
32,
33,
34]. For example, it is not known whether dupilumab, a clinically approved biologic drug that blocks IL4Rα signaling [
35], leads to an in vivo reduction in M2 macrophages in patients receiving this drug. In the future, it would be important to investigate which immune cells are in fact being inhibited by dupilumab. Alternatively, it is possible that dupilumab switches macrophages into a tolerant state, for instance, by turning on the expression of anti-inflammatory molecules. Thus, it is crucial that the mechanism of action of dupilumab be investigated systematically at the cellular and molecular levels. Here, we provide a consideration of macrophages and noncoding RNAs (ncRNAs). Accumulating evidence has revealed that ncRNAs, a class of functional RNAs not translated into proteins and associated with various pathological events, are associated with both macrophage polarization and allergies. ncRNAs are typically classified into two major types that have distinct functions, i.e., housekeeping ncRNAs and regulatory ncRNAs. The detailed classification of ncRNAs is discussed elsewhere [
36,
37]. Emerging evidence shows that these ncRNAs play roles in macrophage polarization related to allergies [
38,
39].
Figure 1. Macrophage polarization. Naïve (M0) macrophages in their inactive state can be polarized into either of two types of activated macrophages with distinct functions, M1 and M2 macrophages (also termed “classically activated” or “alternatively activated” macrophages, respectively), after exposure to certain stimuli. M1 and M2 macrophages are functionally associated with type 1 and type 2 immune reactions, respectively. Several mRNAs and proteins are used as markers to differentiate between these macrophages: i.e., arginase-1 (Arg-1) and CD206 for M2; CD38, CD80, and iNOS for M1 macrophages. However, the criteria for the subclassification of macrophages in vivo in different tissues still require further investigation. Noncoding RNAs might regulate the differentiation of macrophages and/or the function of M1 and M2 macrophages by modifying gene expression programs. Dupilumab is a currently available monoclonal antibody that blocks IL-4 and IL-13 signaling by targeting IL4Rα [
35]. This biologic drug is FDA-approved for allergic diseases such as eczema, asthma, and nasal polyps, which result in chronic sinusitis. Hypothetically, its mechanism of action is in part to inhibit M2 polarization. Tralokinumab, another FDA-approved monoclonal antibody, used for the treatment of atopic dermatitis, targets just the cytokine IL-13 (not depicted). Again, it is not well understood which cell types are being affected by this biologic drug. It would be interesting to directly compare dupilumab versus tralokinumab and assess the in vivo effects of each on macrophages and their noncoding transcriptome. Image created with
BioRender.com (accessed on 28 November 2023).
As mentioned earlier, ncRNAs are RNA molecules that do not encode proteins and are essential in regulating gene expression at both transcriptional and post-transcriptional levels, which includes epigenetic regulation [
60]. There are at least three classes of ncRNAs that regulate gene expression: microRNAs (miRNAs), long ncRNAs (lncRNAs), and circular RNAs (circRNAs). Furthermore, ncRNAs that regulate protein activity have been described [
61,
62]. While miRNA-mediated regulation of gene expression occurs at the post-transcriptional level, lncRNAs and circRNAs may utilize diverse mechanisms of action. Emerging evidence has highlighted the critical role of ncRNAs in regulating macrophage polarization, which may lead to the development of allergies [
63]. Although the mechanisms through which ncRNAs regulate macrophage polarization are diverse and complex, several studies have shown that ncRNAs potentially regulate M1 and M2 macrophage polarization by targeting the regulators of proinflammatory signaling pathways or regulating the expression of anti- or proinflammatory cytokines.
Although published studies have thus far highlighted the functional association between ncRNAs and macrophage polarization or that between ncRNAs and allergic diseases, few reports have described the ncRNA–macrophage polarization–allergy axis. Therefore, in the following sections, we summarize these previous studies on how the individual ncRNA classes are involved in macrophage polarization and how that may relate to allergic diseases.
2. miRNA-Mediated Regulation of Macrophage Polarization
miRNAs are small (typically ~22 nt in length) ncRNAs that post-transcriptionally regulate gene and protein expression by binding to the 3′-untranslated region of the target mRNAs, which induces mRNA degradation and translational repression [
39,
64]. Several miRNAs have been demonstrated to regulate macrophage polarization related to allergic diseases.
The mannose receptor MRC1/CD206 is expressed in immune cells, and its expression level is pronouncedly elevated in M2 macrophages; therefore, it is generally accepted as an M2 macrophage marker [
29]. MRC1/CD206 recognizes an extensive range of surface glycoproteins and plays a crucial role in a variety of immunological events, both physiologically and pathologically [
71]. Interestingly,
miR-511-3p is an miRNA that is transcribed from an intron of the
MRC1 gene. The expressions of
miR-511-3p and MRC1/CD206 have been shown to be coregulated in macrophages [
72,
73]. In studies with the MRC1 knockout mouse model in which
miR-511-3p expression is also deficient, Zhou et al. demonstrated that
miR-511-3p downregulated M1 macrophage polarization, upregulated M2 macrophage polarization, and protected against cockroach allergen-induced lung inflammation [
73]. In addition, it was reported by Do et al. that
miR-511-3p promoted M2 macrophage polarization and attenuated cockroach-allergen-induced lung inflammation by targeting CCL2 [
74]. Alternatively, Heinsbroek et al. demonstrated that
miR-511-3p regulated intestinal inflammation by controlling macrophage-mediated microbial responses via the indirect upregulation of TLR-4 expression [
75]. These findings suggest that
miR-511-3p regulates macrophage functions and polarization by targeting multiple mRNAs.
Using an allergen-induced asthma knockout mouse model, Chung et al. reported that
miR-451a negatively affects IL-4-induced M2 macrophage polarization by targeting and silencing the expression of Sirtuin 2 and promoting asthmatic inflammation [
76]. Additionally, a few other studies were conducted using ovalbumin-induced allergic asthma mouse models to identify miRNAs in macrophage polarization. For example, Veremeyko et al. demonstrated that
miR-124 expression was upregulated in the lung alveolar macrophages of an ovalbumin-induced allergic lung inflammation mouse model and contributed to the development of M2, but not M1, macrophage polarization [
77]. Another study by Shi et al. highlighted the involvement of
miR-142-5p and
miR-130a-3p in pulmonary macrophage polarization and asthma airway remodeling in ovalbumin-sensitized mice [
78]. Additionally, Su et al. reported that
miR-142-5p and
miR-130a-3p functioned by targeting suppressor of cytokine signaling 1 (SOCS1) and peroxisome proliferator-activated receptor γ, respectively [
79]. Notably, this study revealed that SOCS1 had a negative impact on the M2 macrophage polarization in mice [
79], while the M2 polarization of human macrophages is enhanced by SOCS1 [
80]. Again, such contradiction will need to be resolved before ncRNAs can be selected as allergy drug targets.
3. lncRNA-Mediated Regulation of Macrophage Polarization
lncRNAs are long (generally defined to be >200 nt in length) ncRNAs that regulate gene expression at various levels, which include chromatin remodeling, transcriptional regulation, and post-transcriptional regulation [
62,
63]. Several lncRNAs have been shown to regulate M2 macrophage polarization related to allergies. For example, the knockdown of receptor-type tyrosine protein phosphatase ε (PTPRE)-AS1, a lncRNA selectively expressed in IL-4-stimulated macrophages, was shown to promote M2 macrophage activation via the MAPK/ERK 1/2 pathway [
81]. Wen et al. recently demonstrated that MIR222HG acts on the
miR146a-5p/TRAF6/NF-κB axis, leading to the attenuation of macrophage M2 polarization and allergic inflammation in allergic rhinitis [
82]. The few studies investigating the lncRNA AK085865 have also highlighted its role in macrophage polarization [
83,
84]. In particular, the study conducted by Pei et al. showed that AK085865-deficient mice were protected from the allergic airway inflammation induced by Der f 1, a major mite allergen component of
Dermatophagoides farinae [
83]. They also found that AK085865 deletion suppressed M2 macrophage polarization, which subsequently decreased their susceptibility to Der f 1-induced airway inflammation. In addition, Zhang et al. demonstrated that AK085865 specifically interacted with interleukin-enhancer-binding factor (ILF)-2 and functioned as a negative regulator of the ILF2–ILF3-complex-mediated biosynthesis of
miR-192, which promotes M2 macrophage polarization through the direct targeting of interleukin-1 receptor-associated kinase (IRAK) 1 [
84].
lnc-BAZ2B, a lncRNA dominantly expressed in monocytes and significantly upregulated in children with asthma, was also demonstrated to promote M2 macrophage polarization. Mechanistically, lnc-BAZ2B promotes the expression of BAZ2B mRNA by stabilizing its pre-mRNA, leading to enhanced IRF4 expression and M2 macrophage polarization [
85]. Another lncRNA reported to regulate the pathological state of allergies is NKILA [
86]. This lncRNA was demonstrated to limit the asthmatic airway inflammation, enhancing M2 macrophage polarization and inhibiting the NF-κB pathway in a mouse asthmatic model.
In contrast to many reports on the lncRNA-mediated regulation of M2 macrophage polarization in allergy, there are few reports on the lncRNA–M1 macrophage polarization-allergy axis. One of the few such studies, reported by Jiang et al., describes the contribution of lncRNA MEG8-sponging of
miR-181a-5p to M1 macrophage polarization via regulating SHP2 expression in a rat model of IgA purpura, which is a type 3 allergic disease triggered by allergens such as drugs, food, or insect bites [
87]. In another study, Zhu et al. demonstrated that lncRNA growth-arrest-specific transcript 5 (GAS5) is upregulated in exosomes isolated from the nasal mucus of allergic rhinitis patients and promotes M1 macrophage polarization by restraining autophagy and subsequently activating NF-κB signaling [
88].
4. circRNA-Mediated Regulation of Macrophage Polarization
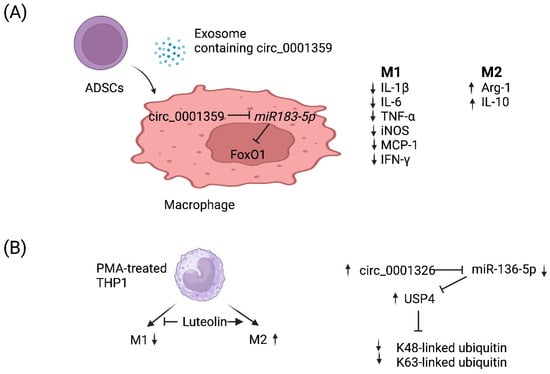
circRNAs are a recently discovered product of back splicing, and a subset of them do not encode for protein. Thus, they comprise a new category of ncRNAs that form covalently closed circular structures, which make them resistant to degradation by RNA exonucleases [
89,
90]. Since they are long-lived, a few circRNAs have been shown to act as molecular “sponges” that sequester miRNAs and/or RNA-binding proteins [
91]. Although the function of most circRNAs remains poorly understood, a few circRNAs have been demonstrated to regulate the macrophage polarization associated with allergy. For example, Shang et al. reported that circ_0001359 was downregulated in ovalbumin-induced asthmatic mice compared with normal mice and circ_0001359-enriched exosomes secreted from adipose-derived stem-cells-attenuated airway remodeling via promoting polarization into M2-like macrophages [
92]. Mechanistically, circ_0001359 was shown to regulate macrophage polarization by enhancing FoxO1 signaling via sponging
miR-183-5p (
Figure 2A) [
92]. Recently, luteolin, a flavone reported to have a protective role in asthma, was shown to activate M2 and suppress M1 macrophage polarization via upregulating circ_0001326 in the human macrophage cell line THP-1 [
93]. The same study also elucidated the underlying mechanism of how circ_0001326 regulates downstream gene expression, including
miR-136-5p and USP4 (
Figure 2B). Finally, it is also conceivable that synthetic circRNAs could be rationally designed to inhibit specific miRNAs to treat diseases such as allergy. Once miRNAs that promote allergy have been identified and validated, then one could simply multimerize the binding sites for miRNA(s) of interest into a synthetic circRNA that will serve to inhibit them and, in turn, allergy.
Figure 2. Working models of how two circRNAs regulate M1 vs. M2 macrophage polarization. (
A) Adipose-derived stem cells (
ADSCs) secrete exosomes that contain circ_0001359. Upon fusion with macrophages, circ_0001359 is released into the cytoplasm and promotes M2-like macrophage polarization in an ovalbumin-induced asthma mouse model and lipopolysaccharide-induced RAW264.7 macrophages cells as evidenced by Arg-1 and IL-10 expression. In contrast, the expression of the following M1 effector molecules is suppressed by circ_0001359: IL-1β, IL-6, TNF-α, iNOS, MCP-1, and IFN-γ. Mechanistically, circ_0001359 inhibits
miR-183-5p via base pairing. Since
Foxo1 mRNA is directly repressed by
miR-183-5p, FoxO1 activity is enhanced as a result and may be in part responsible for reprogramming macrophage cell fate. Image created with
BioRender.com (accessed on 28 November 2023). (
B) Luteolin (a naturally occurring flavonoid found in plants), known for its protective role in asthma, inhibits M1 macrophage polarization and promotes M2 activation in THP-1-derived macrophages. Luteolin-treated THP-1 macrophages induce expression of circ_0001326, inhibiting
miR-136-5p via base pairing. Consequently, ubiquitin-specific protease 4 (USP4) is upregulated since it is directly repressed by
miR-136-5p, and ultimately K48-linked and K63-linked ubiquitin is metabolized by USP4 since it is a deubiquitinase enzyme. Image created with
BioRender.com (accessed on 28 November 2023).
This entry is adapted from the peer-reviewed paper 10.3390/ncrna9060075