Oilseed crops are rich in plant lipids that not only provide essential fatty acids for the human diet but also play important roles as major sources of biofuels and indispensable raw materials for the chemical industry. The regulation of lipid metabolism genes is a major factor affecting oil production.
- oilseed crops
- lipid metabolism
- seed oil content
1. Introduction
Function of Lipids
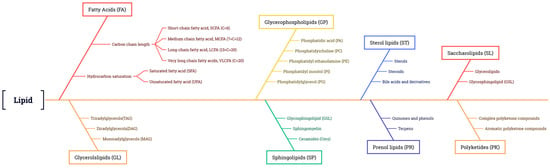
Classification of Plant Lipids
Lipid Accumulation and Storage in Plants
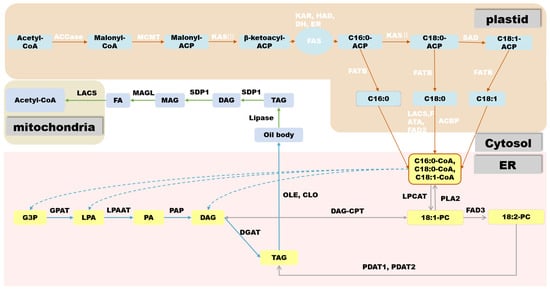
In oilseed crops, the first step in FA biosynthesis is the conversion of acetyl-CoA produced via sucrose glycolysis to malonyl-CoA by acetyl-CoA carboxylase (ACCase). ACCase is a type I biotin-containing enzyme and there are two main forms of ACCase in plants: Heteromeric ACCaseI and Homomeric ACCaseII. ACCase is composed of four subunits: biotin carboxylase (BC), biotin carboxyl carrier protein (BCCP), α-carboxyltransferase (α-CT), and β-carboxyltransferase (β-CT). It has three functional domains, namely the BC functional domain, the BCCP functional domain, and the CT functional domain, respectively [26,27]. After its biosynthesis, malonyl-CoA is transferred to the ACP component of the fatty acid synthase (FAS) complex by malonyl-CoA:ACP malonyltransacylase (MCMT). In plants, FAS is a multi-component type-II enzyme located in the plastids, consisting of 3-β-ketoacyl-ACP synthase III (KASIII), β-ketoacyl-ACP synthase (KASI), ketoacyl-ACP reductase (KAR), hydroxyacyl-ACP reductase (HAD), and enoyl-ACP reductase (ENR). FAS uses acetyl-CoA as the starting unit for a condensation reaction. Each elongation cycle is supplied with a two-carbon unit by malonyl-ACP to produce 16:0-ACP and 18:0-ACP after seven or eight cycles, respectively, at which point 18:0-ACP passes through Δ9 stearoyl-ACP desaturase to form 18:1-ACP, making phosphatidic acid (PA) (16:0) and oleic acid (18:1) the major products of FA biosynthesis in most plant plastids [28].
In most oilseed crops, such as soybean (Glycine max) and rapeseed (Brassica napus), TAGs are stored in seeds as an energy source for germination and aid in seed dispersal [29]. TAG consists of a glycerol backbone and three FA molecules chemically linked by ester bonds, providing a carbon skeleton and energy source. TAG biosynthesis and accumulation occur through a complex network of reactions taking place in the plastid, cytoplasm, and ER. Depending on the plant species, TAG can accumulate in different organs, mainly in embryonic tissues (rapeseed) or endosperm tissues (castor bean). In oilseeds that store oil in the embryo, the main storage tissue is the cotyledons, but substantial seed oil can also accumulate in the hypocotyl, radicle, and surrounding endosperm/aleurone layers [30]. After acyl-CoA is transported from the plastid to the ER, the most common pathway for TAG biosynthesis is the acyl-CoA-dependent Kennedy pathway [31]. In this pathway, acyl-CoA is incorporated into glycerol-3-phosphate (G3P) by acyl-CoA:glycerol-3-phosphate acyltransferase (GPAT) and lysophosphatidic acid acyltransferase (LPAT) at the sn-1 and sn-2 positions of G3P, respectively, to form PA.
Cytoplasmic lipid droplets (LDs) are organelles that store non-polar lipids such as TAGs and sterol esters [33]. In mature seeds, LDs are distributed in the central region of storage cells and are mostly oval or irregular in shape [34]. The primary LD structure consists of a phospholipid monolayer coated with various proteins. The current general model of LD biogenesis is that non-poplar lipids such as TAGs are first produced by membrane-associated enzymes in the ER and then accumulate in the form of a lens between lobes of the ER membrane, culminating with LD formation on the cytoplasmic side of the ER membrane [35]. Two important proteins have recently been shown to be involved in LD formation: SEIPIN [36,37] and lipid-droplet-associated protein in Arabidopsis (Arabidopsis thaliana) [38].
2. Progress in the Identification of Key Genes behind Lipid Metabolism in Plants
3.1. Identification and Functional Characterization of Key Genes
3.2. Transcription Factors Involved in Regulation
3.3. Advances in Multi-Omics Studies of Oilseed Crops
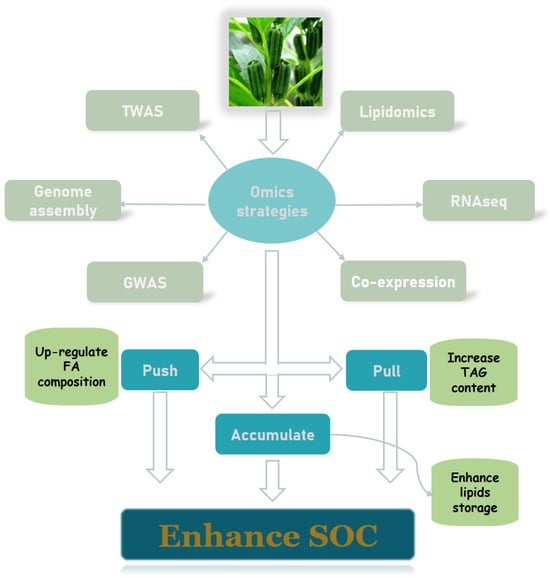
3.3.1. De Novo Genome Sequencing and Annotation
3.3.2. Identification of Differentially Expressed Genes
3.3.3. Construction of Oil Co-Expression Networks
3.3.4. Genome-Wide Association Studies to Map Oil Content–Related Loci
3.3.5. Lipidomics for Oil Structure and Quality Identification
This entry is adapted from the peer-reviewed paper 10.3390/metabo13121170