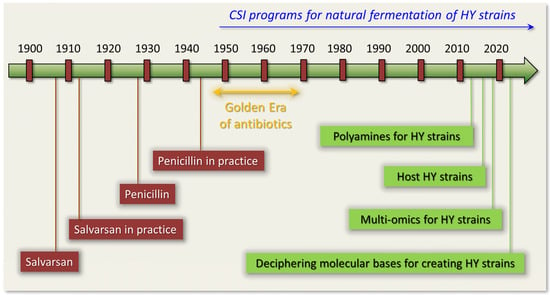
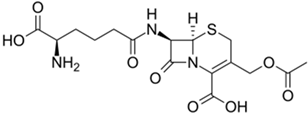
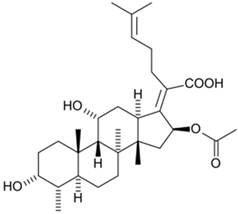
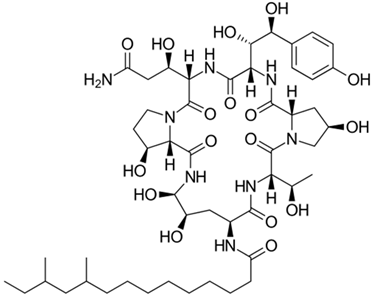
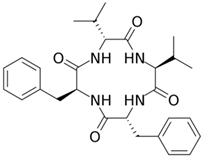
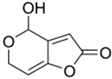
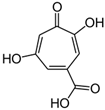
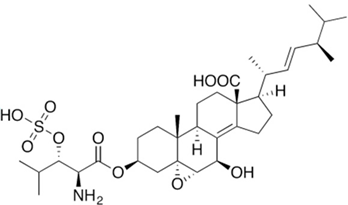
2. Fermentation of Beta-Lactam Antibiotics in Fungi
The fermented penicillin G (PenG) and cephalosporin C (CepC) are modified in vitro to create drugs used in medicine (PenG is also used unmodified) [
152]. Obtaining PenG and CepC in improved fungal strains is more economical than in vitro production [
154]. In fungi, the biosynthesis of these beta-lactams is under the control of a complex hierarchical regulatory system, modified in the process of strain improvement, and is accompanied by a series of biosynthetic reactions, with a significantly increased intensity in HY producers [
94,
155]. In these reactions at the first stage, LLD-ACV tripeptide δ-(L-α-Aminoadipoyl)-L-cysteinyl-D-valine is obtained as a result of non-ribosomal peptide synthesis by the enzyme PcbAB (EC: 6.3.2.26). This tripeptide is then cyclized to isopenicillin N (IPN) through a dioxygenase reaction catalyzed by PcbC (isopenicillin N-synthase (EC: 1.21.3.1)). The IPN core structure is then modified by tailoring the enzyme PenDE (IAT, isopenicillin-N N-acyltransferase [EC:2.3.1.164]) to produce penicillin G, or by sequential reactions of the tailoring enzymes to produce cephalosporin C (CefD1, isopenicillin N-CoA synthetase (EC: 5.1.1.17), CefD2 (isopenicillin N-CoA epimerase (EC: 5.1.1.17), CefEF (penicillin N expandase, EC: 1.14.20.1)/deacetoxycephalosporin C hydroxylase, EC: 1.14.11.26), and CefG (deacetylcephalosporin-C acetyltransferase (EC: 2.3. 1.175)).
2.1. Fermentation of P. chrysogenum for Penicillin G (PenG) Production
The main industrial producer of penicillin G (PenG, or benzylpenicillin, and “Peanut Butter Shot” in military slang) is
P. chrysogenum (formerly known as
Penicillium notatum), which belongs to the class
Eurotiomycetes and the division
Ascomycota [
156,
157,
158]. Currently, a number of molds are known that are capable of producing PenG, for example from the genera
Arthroderma,
Aspergillus,
Penicillium, and
Trichophyton; however, only
P. chrysogenum strains are used for the industrial production of this important antibiotic [
159,
160,
161,
162]. PenG is the first industrially produced natural antibiotic, which was accompanied by an innovative breakthrough in the development of deep fermentation [
163]. From the early 1940s to the present day, PenG has been produced worldwide from the fermentation of improved strains of the
P. chrysogenum NRRL 1951 initial isolate [
79,
164,
165]. This strain was discovered in 1943 growing on a cantaloupe at the local market in Peoria (IL, USA) [
166]. It turned out that
P. chrysogenum NRRL 1951 produces more penicillin than the original Fleming isolate (which produces mostly the unstable and difficult to isolate penicillin F, named after its discoverer) and other
P. chrysogenum industrial strains used at that time for commercial production of PenG (such as
P. chrysogenum NRRL 1249-B21) [
7,
167,
168]. NRRL 1951 was adapted for commercial use; CSI programs were carried out on its basis, which made it possible to increase the PenG yield by more than 1000 times [
41,
101,
169,
170] (
Table 1). Seventy-seven years after its discovery,
P. chrysogenum NRRL 1951 received the status of “State Microbe of Illinois” [
166].
Table 1. Production level of the most important antibiotics from fungi of wild type (WT) and high-yielding (HY) strains derived from them.
Currently, the
P. chrysogenum strains used for PenG production (both the original Fleming strain and the strains used to obtain improved industrial producers) have again been reclassified as
P. rubens [
183]. At the same time, the name
P. chrysogenum was retained for a number of strains that are not used for the industrial production of penicillins and differ from
P. rubens in some characteristics, such as their production of secondary metabolites [
183]. However, in the vast majority of recent publications, it is still common to use the name
P. chrysogenum for PenG producers, in order to avoid confusion [
7,
156]. PenG can be used as a final drug [
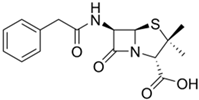
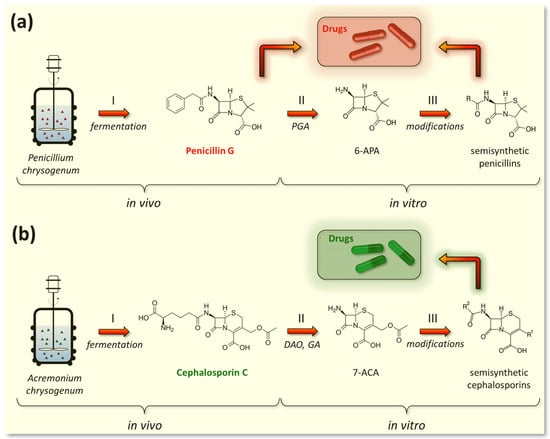
184] or can serve as a raw material for the production of semisynthetic penicillin antibiotics (
Figure 3a). To achieve this, at the first stage, penicillin nucleus or 6-aminopenicillanic acid (6-APA) is obtained from PenG as a result of the deacylation reaction in vitro by amidase enzymes of bacterial origin (penicillin G amidase, EC 3.5.1.11) [
56,
185]. Then, various semisynthetic penicillins are obtained by grafting different side chains onto 6-APA [
186,
187,
188] (
Figure 3a).
Figure 3. Industrial production of beta-lactam antibiotics. (a) Obtaining of penicillin drugs; both penicillin G (a direct fermentation product of Penicillium chrysogenum) and its semisynthetic derivatives are used in pharmaceutical industry. (b) Obtaining of cephalosporin drugs; cephalosporin C (a fermentation product of Acremonium chrysogenum) has low activity and is not used in the pharmaceutical industry, unlike its semisynthetic derivatives. Curly brackets indicate the stages occurring in vivo (stage I—fermentation of fungal strains) and in vitro (stage II—production of core structures and stage III—production of semisynthetic antibiotics). PGA—penicillin G amidase (EC 3.5.1.11); DAO—D-amino acid oxidase (EC 1.4.3.3); GA—glutaryl-7-aminocephalosporanic acid acylase (EC 3.5.1.93); 6-APA—6-aminopenicillanic acid, 7-ACA—7-aminocephalosporanic acid.
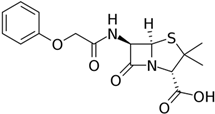
2.2. Fermentation of Penicillins Other Than PenG
Since the 1950s, intensive research has been carried out to produce PenG analogues with altered activity profiles by fermenting
P. chrysogenum with various mono-substituted acetic acids [
189]. As a result, industrial strains of
P. chrysogenum, in addition to benzylpenicillin (PenG), also produce another pharmacologically important compound, phenoxymethylpenicillin (penicillin V, PenV) [
171,
190]. PenG is produced if phenylacetic acid is added to the culture medium as a side chain precursor, and PenV is synthesized if phenoxyacetic acid is added [
185]. To effectively produce PenV in industrial strains, multi-round mutagenesis and selection are also carried out to reduce the level of by-products, such as para-hydroxypenicillin V [
191]. PenV can be used either as a final drug or as a raw material to obtain semisynthetic penicillins through 6-APA (
Figure 3). Currently, only two penicillins produced during fermentation are of industrial importance, namely PenG and PenV [
189]. Other natural penicillins, such as PenF (Fleming’s penicillin), PenK, PenN, PenO, PenU1, PenU6, or PenX, are not currently used in medical practice.
2.3. Fermentation of A. chrysogenum for Cephalosporin C (CefC) Production
The exclusive industrial producer of CefC is
A. chrysogenum (formerly known as
Cephalosporium acremonium), which belongs to the class
Sordariomycetes, division
Ascomycota (
Table 1) [
101]. In contrast to numerous PenG producers, CefC production is found only in a few fungal species. In addition to
A. chrysogenum, the biosynthesis of CefC was also found in
Pochonia chlamydosporia,
Kallichroma tethys, and
Paecilomyces persicinus, where it is not used for industrial production [
159,
162]. This relatively rare distribution of the biosynthetic pathway for this beta-lactam antibiotic in fungi is apparently associated with the appearance of additional genes, the origin of which is discussed in [
7,
192,
193]. In particular, for the biosynthesis of CefC, part of the additional genes is assembled into the so-called “late” beta-lactam BGC, which is located on a different chromosome relative to the localization of the “early” beta-lactam BGC [
89,
193]. Currently, in 2023, the
A. chrysogenum have again been reclassified as
Hapsidospora chrysogena [
100]. Since
Acremonium chrysogenum has been used for a long time to refer the CPC producer, we use this name in current review to avoid confusion. Current industrial producers of CefC are derived from the Brotzu initial isolate
A. chrysogenum ATCC 11550 as a result of CSI programs [
194]. Italian pharmacologist Giuseppe Brotzu isolated
A. chrysogenum ATCC 11550 in 1948 from seawater near Sardinia, Italy [
195]. Industrial descendants of this strain produce several hundred times more CefC [
101,
172,
173]. CefC itself, unlike PenG, has weak antibacterial activity and is not used as a final drug in medical practice (
Figure 3b) [
196]. The key role of cephalosporin C is associated with its use as a starting substance for the production of all pharmaceutically important cephalosporin antibiotics [
197]. To achieve this, at the first stage, cephalosporin nucleus or 7-aminocephalosporanic acid (7-ACA) is obtained in vitro from CefC, either chemically or enzymatically (using D-amino acid oxidase (EC 1.4.3.3) and glutaryl-7-aminocephalosporanic acid acylase (EC 3.5.1.93)) [
56,
198,
199]. The side groups of 7-ACA are then modified by various chemical or enzymatic methods to produce cephalosporin drugs [
56,
169,
200,
201].
3. Fermentation of Non-Beta-Lactam Antibiotics in Fungi
The fermentation of fungal strains is used to produce a number of non-beta-lactam antibiotics for medical use. Some of them, such as fusidic acid and griseofulvin, were introduced to the market during the Golden Age of antibiotics and are still used today [
202,
203]. Other antibiotics, such as pleuromutilins, were discovered in the early 1950s, but only entered the market as semi-synthetic derivatives in 2007 [
204,
205,
206]. The emergence of new classes of antibiotics from fungi, which entered the market in the last 20–25 years, was one of the most important factors that made it possible to fill the innovative gap in the introduction of antibiotics that arose after the end of the Golden Era of antibiotics [
65]. For example, relatively recently discovered antibiotics, from the classes echinocandins and enfumafungins, were introduced into medical practice at the beginning of the 21st century [
207,
208]. There are also antibiotics that were previously obtained using fermentation in fungal strains, but that have now been discontinued due to discovered toxicity. For example, fusafungine, which belongs to the enniatin class of antibiotics, was withdrawn from the market in 2016 due to toxicity [
209]. Although this review focuses only on antibiotics produced by fungal fermentation, it should be noted that improved bacterial strains are also widely used for the industrial production of non-beta-lactam antibiotics [
210,
211,
212]. For example, some major market antibiotics, such as macrolides and aminoglycosides, and some minor market antibiotics, such as tetracyclines and glycopeptides, were originally isolated from
actinomycetes, especially from the genus
Streptomyces (
Figure 2) [
210]. Bacteria are also a promising source for obtaining new antibiotics, for example, antimicrobial peptides [
213]. On the other hand, there are vast classes of non-beta-lactam antibiotics, such as fluoroquinolones and sulfonamides, which are obtained by chemical synthesis [
214,
215].
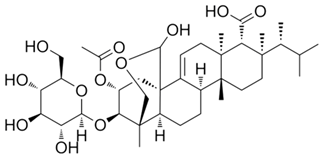
3.1. Fusidanes
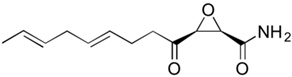
Steroid antibiotics from the fusidane class, such as fusidic acid, helvolic acid, and cephalosporin P1, are produced by various filamentous fungi [
153]. Although these antibiotics have been known since the 1960s, they have drawn renewed attention, due to the fact that they have no cross-resistance to generally used antibiotics [
174,
216]. Fusidic acid, the key fusidane antibiotic, is obtained from the fermentation of improved strains of
Fusidium coccineum [
176]. The production level in the improvement process was increased by a hundred times (
Table 1). Other fusidanes, helvolic acid from
Aspergillus fumigatus and cephalosporin P1 from
A. chrysogenum, do not currently have such extensive use in medicine (
Table 2) [
217,
218].
Table 2. Some examples of antibiotic production by fungal strains.
3.2. Griseofulvin
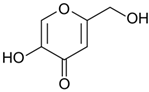
Griseofulvin, an ascomycete polyketide metabolite that interferes with the polymerization of fungal microtubules, was marketed in 1959 [
230]. It was first isolated in 1939 and is still widely used in fields ranging from medicine to agriculture [
230,
231]. The industrial fermentation of griseofulvin is carried out on the basis of improved strains of
Penicillium griseofulvum (reclassified from
Penicillium patulum) [
84,
232]. The production level is several tens of grams per liter; however, specific numbers and methods of classical improvement are described in the relevant patents and are successfully masked [
179,
230].
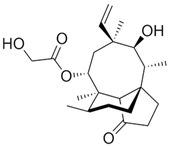
3.3. Pleuromutilins
Pleuromutilins are SMs of the terpene type [
24]. Despite the fact that pleuromutilin, the first representative of this class of antibiotics, was discovered back in 1951, pleuromutilins only recently entered the market as a medicine for humans [
204]; retapamulin, the first antibiotic of this class, which is approved for humans, began to be used in 2007 [
233,
234]. To obtain the semisynthetic antibiotic retapamulin, pleuromutilin is used, which is produced as a result of fermentation of
Clitophilus scyphoides and other basidiomycetes [
205,
235]. In 2019, lefamulin, another semisynthetic antibiotic from the pleuromutilin class, was approved for systemic use in humans [
209,
236,
237,
238]. An important characteristic of pleuromutilins is the lack of cross-resistance with other antibiotics, such as the macrolides, which act on a similar target, the 50S subunit of the prokaryotic ribosome [
239].
3.4. Echinocandins
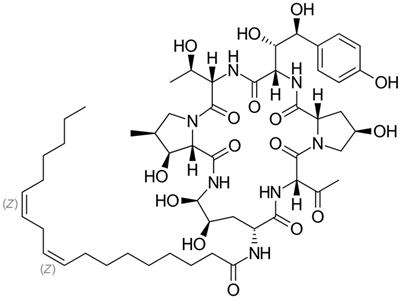
Currently, more than 20 echinocandins, lipopeptide antibiotics, have been isolated from fungi, the fermentation of three of which—echinocandin B, pneumocandin B
0, and FR901379—has been commercialized [
240]. They are used as raw materials in semisynthetic synthesis to obtain anidulafungin, caspofungin, and micafungin, respectively, which were introduced into medical practice in the 2000s [
207,
241,
242,
243]. These compounds are also called “antifungal penicillins” because they act on the biosynthesis of the cell wall of fungi, just as penicillins destroy the cell wall of bacteria [
244]. Echinocandin B was first isolated in 1974 from
A. nidulans, becoming the first antibiotic belonging to the echinocandin class [
245]. Currently, it is produced by improved strains of
A. nidulans and is used as a raw material for the semisynthetic antibiotic anidulafungin, used in medical practice [
246,
247]. Industrial strains of
A. nidulans produce approximately a thousand times more echinocandin B than WT strains [
181]. Caspofungin, another semisynthetic lipopeptide widely used in medicine, is derived from pneumocandin B
0, which is produced by fermentation of the ascomycete
Glarea lozoyensis [
248]. Wild-type strains of
G. lozoyensis predominantly synthesize pneumocandin A
0, approximately 10 times more than pneumocandin B
0 [
240]. As a result of CSI programs, HY producers of pneumocandin B
0 were obtained, which in its structure is less similar to echinocandin B than pneumocandin A
0 [
180,
249,
250,
251]. Now, industrial strains produce hundreds of times more pneumocandin B
0 compared to WT strains [
180]. One of the most important antibiotics of this class, semisynthetic micafungin, is derived from the natural product FR901379 (via the enzymatic synthesis of compound FR179642) [
252,
253]. Lipopeptide FR901379 is obtained as a result of the natural fermentation of the improved strains of the ascomycete
Coleophoma empetri [
254,
255]. Recently, the industrial producer
C. empetri MEFC009, using a genetic engineering approach, was able to further increase the yield of FR901379 [
182,
256].
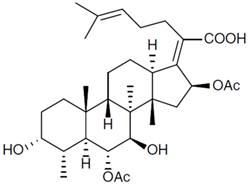
3.5. Enfumafungins
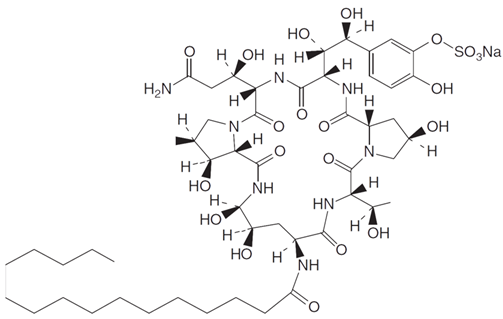
Enfumafungin is a glycosylated triterpenoid that acts on the fungal cell wall [
257]. This SM, like the echinocandins, targets β-1,3-glucan synthase, but appears to bind this multiprotein complex at a different site [
258]. Due to this, there is a lack of cross-resistance between these two classes of antibiotics, and pathogenic fungi that have acquired resistance to echinocandins show sensitivity to enfumafungins [
259,
260]. Enfumafungin is obtained from the ascomycete
Hormonema carpetanum [
220]. In 2021, ibrexafungerp, a semi-synthetic derivative of enfumafungin, was introduced into medical practice, representing the first non-azole oral antifungal drug to be approved in the United States for the treatment of vaginal yeast infections [
208,
261] (
Table 2).
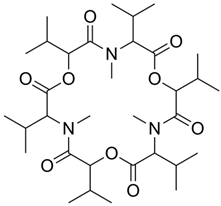
3.6. Enniatins
Enniatins are cyclohexadepsipeptides that are produced by
Fusarium,
Verticillium,
Halosarpheia, and several other fungal genera [
262]. Currently, about thirty natural compounds of this class have been characterized [
219]. The first enniatins were discovered more than 70 years ago [
263]. Since then, the antibiotic fusafungine has been introduced into clinical practice [
264]. It is a cocktail of enniatins resulting from the fermentation of
Fusarium lateritium strains, due to partially the non-specific work of non-ribosomal multifunctional enzyme enniatin synthetase (NRPS) [
219,
265,
266,
267]. The European Medicines Agency, in 2016, recommended the withdrawal of fusafungine from the market due to possible rare but severe allergic reactions [
209,
268].
4. Fungal Secondary Metabolites for the Development of Novel Drugs
Fungi produce significant amounts of SMs with antimicrobial properties [
71,
269]. Only a few of them have been introduced into medical practice (
Table 1 and
Table 2) [
71,
203]. However, there are a significant number of compounds that have demonstrated antibiotic or other medically important activity [
23]. A case in point is a literature survey showing that, among 1500 compounds isolated from fungi in the 1990s, more than half exhibited antibacterial, antifungal, or antitumor activity [
270]. Modern approaches that combine advances in the fields of genetic engineering and bioinformatics have made it possible to significantly increase the number of fungal SMs that are potentially significant for human use [
271,
272]. Some of these compounds and their semisynthetic derivatives are currently in clinical trials at phase I or greater [
71,
273]. Thus, the semisynthetic beta-lactam antibiotics benapenem, sulopenem, and its prodrug sulopenem etzadroxil, which act on the cell wall of bacteria, are in phase-III clinical trials [
273,
274,
275,
276]. Semisynthetic beta-lactams sanfetrinem and its prodrug sanfetrinem cilexetil are in phase-II clinical trials [
273,
277,
278]. Several fungal antibiotics are currently being tested in clinical trials for the treatment of other diseases, such as cancer. For example, wortmannin, an antifungal furanosteroid antibiotic isolated in 1975 from
Talaromyces wortmannii (formerly
Penicillium wortmannii) [
279], was then also shown to have inhibitory activity against phosphatidylinositol 3-kinase [
280]. Currently, a semisynthetic compound PX-866 has been developed based on wortmannin, which has entered phase II clinical trials for recurrent glioblastoma [
281]. Fungal SMs are being tested in clinical trials for the treatment of a wide range of diseases [
71]. For example, muscimol, an isoxazole from
Amanita pantherina, is an ionotropic GABA
A-R receptor agonist and was developed for the treatment of drug-resistant epilepsy (phase I) [
282]. Psilocybin from hallucinogenic fungus
Psilocybe mexicana binds to the 5-HT
2A serotonergic receptor and is developed for treatment-resistant depression [
283,
284]. Antroquinonol was isolated in 2007 from
Taiwanofungus camphoratus (formerly
Antrodia camphorata), a parasitic fungus indigenous to Taiwan [
285]. It inhibits isoprenyl transferase, an enzyme that is involved in the post-translational processing of Ras and Rho proteins, which leads to the inhibition of Ras and Rho signaling, and that has antiviral, anti-inflammatory, anti-fibrotic, anti-cancer, and anti-hyperlipidemia properties [
286]. This ubiquinone derivative was entered into phase II clinical trials for treating non-small-cell lung cancer [
287]. The introduction, in recent years, of new classes of antibiotics from fungi (and their semi-synthetic derivatives), such as echinocandins, enfumafungins, and pleuromutilins, indicates significant potential for the development of novel drugs based on fungal SMs.