The presence of lipoarabinomannan (LAM) in the Mycobacterium tuberculosis (Mtb) cell envelope was first reported close to 100 years ago. Since then, numerous studies have been dedicated to the isolation, purification, structural definition, and elucidation of the biological properties of Mtb LAM. The significance of LAM remains high to this date, mainly due to its distinct immunological properties in conjunction with its role as a biomarker for diagnostic tests due to its identification in urine, and thus can serve as a point-of-care diagnostic test for tuberculosis (TB). LAM has been thoroughly studied and massive amounts of information on this intriguing molecule are now available.
1. Introduction
The hallmark of
Mycobacterium tuberculosis (
Mtb) is a complex hydrophobic lipid-rich cell envelope [
1]. The dominant features have been studied by electron microscopy [
2] and biochemical fractionation studies [
3]. The
Mtb cell envelope is mainly composed of four distinct entities [
3]. Surrounding the cytosol is the cytoplasmic membrane or innermost membrane, which is similar to other bacterial membranes in thickness and mainly composed of proteins and phospholipids such as phosphatidic acid (PA), diphosphatidylglycerol (PG), phosphatidylcholine (PC), phosphatidylethanolamine (PE), and the phosphatidyl-
myo-inositol mannosides (PIMs). The cell envelope lipoarabinomannan (LAM) and lipomannan (LM) are lipoglycans found anchored in the plasma membrane via a phosphatidyl-
myo-inositol moiety that is the structural basis of the PIMs. Beyond the cell membrane, peptidoglycan (PG) and its attached arabinogalactan (AG), which together form the AGP complex (AGP), are apparently located in the periplasmic space. Outermost is an outer membrane or “
mycomembrane”, which consists of the mycolic acids that form a distinct layer [
4]. This peripheral lipid-rich cell envelope and its distinct components have been defined as virulent factors due to their ability to trigger a host response detrimental to the host and favoring the establishment of an
Mtb infection. A debate still exists on the exact location of the PIMs, LM, and LAM. The latter may be anchored in the plasma membrane during its biosynthesis but also exposed on the cell envelope surface. Evidence of the existence of “
kettle holes” on the cell envelope poles, which could allow the surface exposure of plasma membrane anchored LAM [
5,
6,
7,
8], could explain this seeming contradiction. The concept of the exposure of LAM on the surface is supported by quantitative transmission electron microscopy studies indicating that LAM is mainly located in the bacillus poles/tips and within cell envelope surface depressions [
9,
10,
11]. Meanwhile, the concept of LAM anchored to the plasma membrane is supported by the need for mechanical and chemical steps to extract LAM from the bacterial cell envelope [
12] and by the fact that many steps in the LAM biosynthesis are associated with the plasma membrane [
13,
14].
2. Defining the Basic Structure of LAM
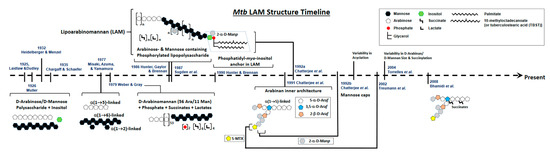
A timeline of the evolution of LAM structural studies is presented in
Figure 1. Briefly, in 1925, Laidlaw and Dudley [
15] used a mild alkaline extraction of a defatted human strain of
Mtb to obtain a novel component that contained 31% pentose. Later, Heidelberger and Menzel using dilute acetic acid on defatted
Mtb [
16], as well as Laidlaw and Dudley and Mueller using culture filtrate, showed the presence of a serologically active polysaccharide rich in D-arabinose and D-mannose [
17]. The presence of this polysaccharide was confirmed further in avian and bovine strains [
18]. Over the next 10 years, several studies followed, all concluding the presence of a certain polysaccharide composed of D-mannose, D-arabinose, and an unidentified sugar acid that reacted as haptens.
Figure 1. Schematic Representation of the evolution of Mtb LAM structural studies. Approximate timeline for last 100 years starting in 1925.
The potential first evidence of a mycobacterial LAM-like structure was established in 1935 by Chargaff and Schaefer, who identified the polysaccharide, designated as fraction A, as being a soluble, dextrorotatory, weak acid containing 77% reducing sugars and 3% amino sugars, mainly composed of mannose and arabinose together with small amounts of inositol [
19].
In 1977, a serologically active and LAM-related D-arabino-D-mannan from
Mtb was characterized with a 2:1 ratio of arabinose:mannose. This arabinomannan consisted of α-(1 → 5)-linked D-arabinose residues and α-(1 → 6)- and (1 → 2)-linked D-mannose residues. Further methylation and enzymatic degradation studies using
Arthrobacter sp. α-D-mannosidase and M-2 enzyme (D-arabinan hydrolase) provided evidence of the existence of short side chains built up from α-(1 → 2)-D-mannosidic linkages that were attached to an α-(1 → 6)-linked mannan backbone [
20].
In 1979, Weber and Gray [
21] isolated an acidic arabinomannan from
M. smegmatis and defined the presence of 56 arabinosyl and 11 mannosyl residues, 2 phosphates, 6 mono-esterified succinates, and 4 ether-linked lactate groups. Subsequently, this acidic polysaccharide was separated into phosphorylated and non-phosphorylated forms with similar structures, wherein the main structural feature was the presence of chains of contiguous arabinofuranosyl residues linked α-(1 → 5).
Close to 50 years since the first reported evidence of a polysaccharide containing arabinose, mannose, inositol, and phosphorous in
Mtb, in 1986, Hunter et al. structurally defined a family of major arabinose- and mannose-containing phosphorylated lipopolysaccharides isolated from
Mtb and
M. leprae [
22]. LAM-B, as designated, eluting off an ion exchange column, in addition to arabinose and mannose, also contained glycerol and a
myo-inositol 1-phosphate, as well as acylations with lactate, succinate, palmitate, and 10-methyloctadecanoate. Other studies followed that provided details of the arabinose-to-mannose ratios and the presence of palmitic, stearic, and tuberculostearic acids as the main fatty acids [
23].
Chatterjee and colleagues, using enzymatic treatments, subsequently focused on the exact architecture of the arabinan domain, establishing that its internal regions consist of branched 3,5-linked α-D-Ara
f units with stretches of linear 5-linked α-D-Ara
f residues attached at both branch positions, whereas the non-reducing terminal segments of the LAM arabinan domain consist of either of the two arrangements, Ara
4: β-D-Ara
f-(1 → 2)-α-D-Ara
f-(1 → 5)-α-D-Ara
f-(1 → 5)-α-D-Ara
f → or Ara
6: [β-D-Ara
f-(1 → 2)-α-D-Ara
f-(1 → ]
2 → (3 and 5)-α-D-Ara
f-(1 → 5)-α-D-Ara
f → [
27] (
Figure 2). Two subsequent studies by Chatterjee and colleagues further refined the LAM structure. The first focused on describing the structural environment of the PI anchor, showing clearly the presence of the 1-(sn-glycerol-3-phospho)-D-
myo-inositol-2,6-bis-α-D-mannoside unit, indistinguishable from that derived from phosphatidyl-
myo-inositol dimannoside (PIM
2). The same study also demonstrated that the C-6 position of inositol is the site of attachment of the mannan core of LAM, which consists of an α(1 → 6)-linked backbone with several α(1 → 2)-side chains [
28]. In a parallel study, Chatterjee and colleagues also demonstrated that the termini of LAM from virulent
Mtb strains, unlike those from attenuated
Mtb strains, are extensively capped with mannosyl (Man
p) residues, either with a single α-D-Man
p, a dimannoside (α-D-Man
p-(1 → 2)-α-D-Man
p) or a trimannoside (α-D-Man
p-(1 → 2)-α-D-Man
p-(1 → 2)-α-D-Man
p). Thereby, the functionally important so-called mannose-capped LAM or Man-LAM was identified (
Figure 3).
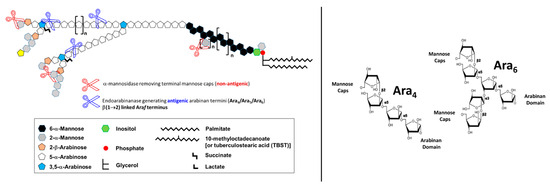
Figure 2. Schematic representation of ManLAM from
Mtb. The antiLAM monoclonal antibodies (mAbs) CS-35, A194-01, and CHCS9-08 are shown to react with the LAM arabinan terminal arrangements (Ara
4 and Ara
6, respectively). These three mAbs are widely used in immunoassays for TB diagnosis. The cartoon is based on a screening of 12 synthetic arabinan glycoconjugates by Dr. Todd Lowary [
40]. In order to identify which motifs of LAM are being recognized by anti-LAM antibodies, an exhaustive digestion of LAM with commercially available α-mannosidase (derived from Jack Beans) is performed. This digestion removes the mannose caps of LAM (depicted as grey hexagons). An additional digestion using an endo-arabinanase (in-house isolated from
Cellulomonas) releases arabinan fragments from the D-arabinan domain, mainly Ara
4, Ara
5, and Ara
6 fragments. These enzymatic digestions allowed the identification of the Ara
4 and Ara
6 motifs as the ones being recognized by monoclonal antibodies against LAM and were used in the development of TB diagnosis downstream.
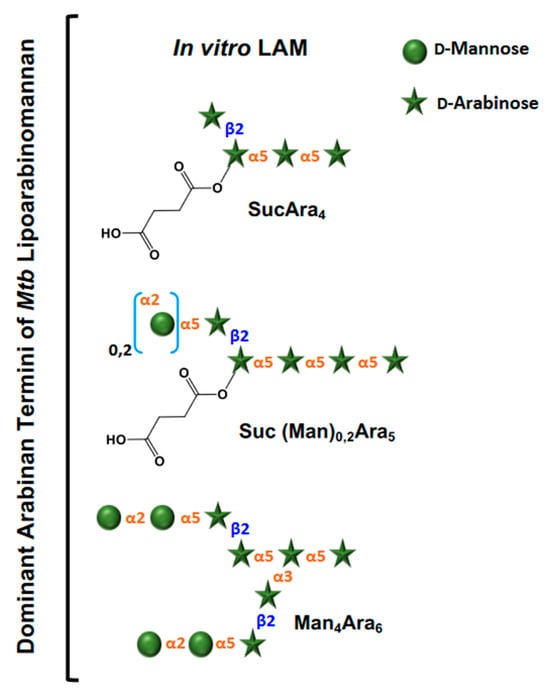
Figure 3. Dominant arabinan termini of
Mtb lipoarabinomannan from culture. Only non-reducing ends are shown. The terminal structures evolved after extensive enzymatic degradation of LAM followed by liquid chromatography with tandem mass spectrometry (LC/MS-MS) analyses [
41]. Abbreviations: Ara = D-Arabiniose, Man = D-Mannose, Suc = Succinates.
3. Evolution of LAM Structural Studies across Species
Essentially, LAM throughout contains four structural domains: a mannosylated phosphatidyl myo-inositol (PI) anchor, a D-mannan core, a D-arabinan domain, and different capping motifs that contribute to species and strain diversity [43,44]. The mannan core consists of a chain of α-(1 → 6)-linked mannopyranose (Manp) residues, some of which are modified by the addition of α-(1 → 2)-linked Manp motifs, usually, but not always, as a single residue. An arabinan, composed of solely D-arabinofuranose (Araf) residues, is attached to the non-reducing end of the mannan core [42]. Capping motifs can be added at specific positions contributing to intra- and inter-species structural variability [41]. Fast-growing mycobacterial species predominately produce AraLAM (uncapped LAM) or PILAM (phosphoinositol capped, as defined for M. smegmatis) [45]. Slow-growing mycobacteria like Mtb and M. leprae produce LAM with α-(1 → 2)-linked Manp capping residues, giving a molecule referred to as ManLAM [8,30]. Within the Mtb complex group, variations regarding primarily the degree of terminal mannose capping present in ManLAM can range between 40–70% [27,28,46,47,48]. Some fast-growing and/or non-pathogenic mycobacteria also produce ManLAM; however, these differ in the Man content of the capping motifs. In addition to Manp capping, ManLAM from strains of the Mtb complex group also contains a unique residue –MTX on the terminal Manp caps [34,35,36,49].
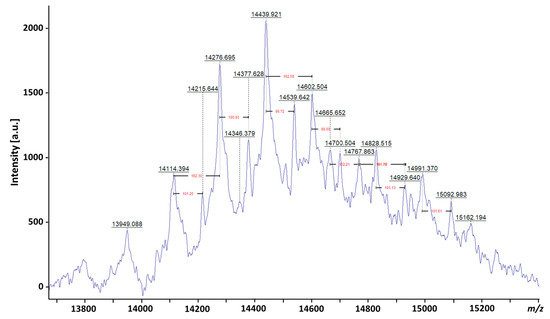
The extreme heterogeneity in LAM is evident from the broad diffuse band observed on a SDS-polyacrylamide gel electrophoresis (PAGE) analysis of LAM and LM [
22] and its capacity to be separated in different isoforms [
31], as well as from several recent matrix-assisted laser desorption/ionization—mass spectrometry (MALDI-MS) studies, which provided an indication of the mean distribution of true molecular mass. It has been shown that native LAM from
M. bovis BCG and
Mtb gives a broad peak centered at 17.3 kDa before deacylation and 16.7 kDa after deacylation, with a reported size distribution range of ±4 kDa depending on the studied strains [
52]. With recent advances in MS instrumentation, a peak centered at
m/
z 14439.921, providing a molecular mass of approx. 15 kDa for LAM, has been reported (Chatterjee et al., personal communication) (
Figure 4).
Figure 4. MALDI-TOF analysis of LAM from
Mtb. The mass spectrometry (MS) was done on Bruker ultrafleXtreme matrix-assisted laser desorption time-of-flight spectrometry (MALDI-TOF/TOF MS), indicating a molecular mass for ManLAM of approximately 15 kDa. Heterogeneity in mass of 162
m/
z corresponds to a hexose and 100
m/
z corresponds to succinates.
4. Biological Properties of Mtb LAM
Mtb LAM was widely studied in the 1990s. Studies started to reveal the immunological properties of LAM, initially as a potential candidate for ELISA-based diagnosis of TB and leprosy [
53,
54], as well as its capacity to inhibit the activation of macrophages, among other immunological features [
55,
56].
Early data published include LAM-induced abrogation of T-cell activation [
57]; inhibition of various IFN-γ-induced functions including macrophage microbicidal and tumoricidal activity [
56]; scavenging of potentially cytotoxic oxygen free radicals [
58]; inhibition of protein kinase C activity [
58]; and evocation of a large array of cytokines associated with macrophages such as TNF [
59,
60,
61,
62,
63], GM-CSF, IL-1α, IL-1β, IL-6, and IL-10 [
64,
65]. In 1991, Chan and colleagues provided the first evidence of the role of
Mtb LAM in downregulating macrophage effector functions by scavenging potentially cytotoxic oxygen free radicals, inhibiting protein kinase C activity, and blocking the transcriptional activation of IFN-γ inducible genes [
58]. Related to the use of LAM by
Mtb to recognize and infect host cells, it has been reported that both the mannose receptor (MR) and DC-SIGN in phagocytes recognize and bind to LAM, and this binding can result in efficient internalization of
Mtb to its intracellular niche within host cells [
9,
66].
Of the many biological properties that
Mtb ManLAM has exhibited, one has led us to understand how critical this molecule is for the persistence of
Mtb within host cells. First, Schlesinger and colleagues showed that ManLAM is recognized by macrophages [
67]. This finding was later supported by others [
68]. Subsequently, Deretic and others showed the capacity of
Mtb ManLAM to block phagosome acidification [
69], and later, Schlesinger and colleagues showed how
Mtb exploits the ManLAM/MR route [
70] to gain entry into macrophages and survive [
9], potentially generating the perfect niche for survival and subsequent immune responses deriving into granuloma formation and thus, contributing to the persistence of
Mtb in a latent stage in the host.
5. LAM as a Diagnosis Biomarker for TB Disease
Mtb LAM represents up to 15% of the bacterial mass [
1,
26,
74,
75]. This molecule is firmly but non-covalently attached to the inner membrane and extends to the exterior of the cell envelope [
76], where it interacts as a potent virulence factor modulating host immune responses [
77]. Importantly, the linear terminus Ara
4: (β-D-Ara
f-(1 → 2)-α-D-Ara
f-(1→5)-α-D-Ara
f-(1 → 5)-α-D-Ara
f) (
Figure 2) and a branched terminus Ara
6: ([β-D-Ara
f-(1→2)-α-D-Ara
f-(1-)
2 → 3, and →5]-α-D-Ara
f-(1→5)-α-D-Ara
f) (
Figure 2) are shown to be the epitopes recognized by anti-LAM monoclonal antibodies (mAb) [
40,
74,
78,
79,
80,
81].
LAM has been validated to be present in variable concentrations in sputum, serum, and urine [
82,
83,
84]. In recent years, several laboratories including ours have made significant advances toward developing urinary LAM-based diagnostics for active TB [
40,
79,
85,
86,
87]. A point-of-care (POC) test that readily diagnoses active TB would reduce diagnostic delays, interrupt transmission with appropriate therapy, and address many of the current gaps in global TB control (Stop TB Partnership in collaboration with Imperial College London). The development of sensitive POC methods to detect LAM in non-invasive samples such as urine using immunoassays is currently stagnant due to the suboptimal sensitivity of the assays. Current methods are also limited in applicability to TB diagnosis only in people living with HIV and severe disease [
84,
88,
89]. However, there are studies showing that LAM can be also detected in urine from active TB cases without HIV co-infection [
85,
90]. Recently, as an alternative to urine, groups have focused their attention on other non-invasive samples such as exhaled breath condensate (EBC), in which ManLAM seems to be present in significant amounts [
91,
92].
This entry is adapted from the peer-reviewed paper 10.3390/pathogens12111281