Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Subjects:
Behavioral Sciences
Sequence learning is a fundamental ability of the human brain. It forms the basis of many cognitive, social, and motor skills. Repetitive transcranial magnetic stimulation (rTMS) is an increasingly used non-invasive brain stimulation (NIBS) tool to examine the functional role of cortical areas and brain networks. In addition to neuroimaging methods, rTMS might contribute to a better understanding of the functional and neural underpinnings of visuomotor sequence learning.
- non-invasive brain stimulation
- sequence learning
- repetitive TMS
1. Stimulated Brain Regions
First, the researchers examined the stimulated brain regions and found that the two most frequent targets were M1 (nine out of 17 studies) and the DLPFC (eight out of 17 studies). In addition, three studies targeted the SMA and four the parietal cortex. One study stimulated the Broca area, and another one the cerebellum (see Figure 3).
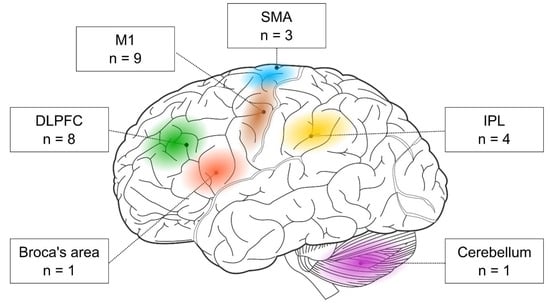
Figure 3. The cortical targets of rTMS-SRTT studies. Brain image was adapted from Hugh Guiney (https://commons.wikimedia.org/wiki/File:Human-brain.SVG; CC BY-SA 3.0; accessed on 25 July 2021).
M1. The role of M1 in motor consolidation goes beyond sequence learning as it engages in the early motor consolidation of elementary motor behavior (Buetefisch et al. 2015; Bütefisch et al. 2004; Muellbacher et al. 2002). The popularity of M1 as a stimulation target is attributed to its implicated role in the initial encoding of sequences and the early consolidation of already learned sequences (Seidler et al. 2005). In the reviewed rTMS studies, typically, low-frequency rTMS or cTBS was applied over M1, resulting in the weakening of the learning process (Clark et al. 2019; Rosenthal et al. 2009; Steel et al. 2016; Wilkinson et al. 2015), or the prevention of offline improvements (Breton and Robertson 2017; Robertson et al. 2005). Interestingly, two studies found an increase in visuomotor skills following low-frequency rTMS or cTBS over M1. In one of them, low-frequency rTMS affected the SRTT indirectly through the prevention of interference with a declarative task (Cohen and Robertson 2011). In the other study, cTBS abolished the decrease in corticospinal excitability, which allowed for offline improvements on an explicit SRTT (Tunovic et al. 2014). Only one of the nine studies attempted to use iTBS over M1, but it did not find any effect on implicit sequence learning (Wilkinson et al. 2010).
DLPFC. The DLPFC has traditionally been identified as a brain area supporting executive functions and working memory (Miller and Cohen 2001; Yuan and Raz 2014). Plasticity changes in the DLPFC seem to be associated with sequence learning (Cao et al. 2022). Lesion studies—where patients with prefrontal lesions show decreased sequence learning on the SRTT (Beldarrain et al. 1999, 2002)—highlight its importance in visuomotor sequence learning, too. However, its functional role is still controversial (Janacsek and Nemeth 2013, 2015). Based on recent models, the DLPFC may act as a neural switch between competitive memory processes (Ambrus et al. 2020; Daw et al. 2005; Lee et al. 2014). On the one hand, it may favor declarative learning and memory (e.g., memory for events and facts), as well as top-down processes. However, if the situation requires acquiring new regularities (e.g., a completely new pattern or sequence), it recedes. A potential mediator role of the DLPFC is also supported by the findings of a TBS study, where the learning of linguistic sequences was enhanced due to the disruptive stimulation of the DLPFC (Smalle et al. 2022). Out of the eight identified studies, three studies found that rTMS over the DLPFC weakened implicit sequence learning (Cohen and Robertson 2011; Pascual-Leone et al. 1996; Robertson et al. 2001). In one of them, DLPFC stimulation reduced learning on the SRTT indirectly by interfering with a declarative task (Cohen and Robertson 2011). Examining explicit sequence learning, two of the six studies found an enhancement for cTBS over the DLPFC (Galea et al. 2010; Tunovic et al. 2014). However, one recent study found no effect of DLPFC stimulation on explicit the SRTT (Gann et al. 2021). Two additional studies used probabilistic instead of deterministic sequences. One of them found that low-frequency rTMS over the DLPFC led to better performance on this sequence type (Ambrus et al. 2020), while the other study found no effect of DLPFC stimulation on performance (Wilkinson et al. 2010).
SMA. Some neuroimaging studies reveal that, besides M1, another motor area, the SMA also appears to be involved during the SRTT (Hazeltine et al. 1997; Seidler et al. 2005). According to an fMRI study, SMA activation is associated with the performance of sequential movements (Hikosaka et al. 1996). Additionally, a PET study suggests that the SMA is involved in the execution of previously learned sequences rather than in the acquisition of sequences (Honda et al. 1998). In more recent studies, the SMA was found to be involved in the automatization of sequential movements (Shimizu et al. 2020) and the consolidation of implicit sequence knowledge (Verwey et al. 2022).
Parietal cortex. According to previous behavioral studies, a motor response is not strictly necessary for the acquisition of complex sequences; monitoring in itself can lead to learning (Nemeth et al. 2009; Song et al. 2008; Zolnai et al. 2022). Based on functional neuroimaging studies, the inferior parietal lobule (IPL) encodes the sequence at a general, abstract level, independently of the response mode (Grafton et al. 1998; Hikosaka et al. 1999).
Broca’s area. In the field of sequence learning, Broca’s area has been primarily tested on artificial grammar learning tasks (De Vries et al. 2010; Uddén et al. 2017), where participants need to extract rules from artificially generated grammatical sequences (Reber 1967, 1989). Because the acquisition of the grammar of a language is connected to sequence learning (Nemeth et al. 2011), it is an interesting question whether this brain area is also involved in the acquisition of non-linguistic visuomotor sequences. Only one study has examined the role of Broca’s area and showed that cTBS over the BA 44 prevented the learning on an implicit SRTT (Clerget et al. 2012).
Cerebellum. The role of the cerebellum in sequence learning is highly uncertain in the literature (Baetens et al. 2020; Janacsek et al. 2020). While the detrimental effect of cerebellar damage on sequence learning assumes its essential role (Dirnberger et al. 2013; Doyon et al. 1997; Gomez-Beldarrain et al. 1998; Shin and Ivry 2003), neuroimaging studies do not always support this hypothesis (Janacsek et al. 2020; Kóbor et al. 2022; Seidler et al. 2002; van der Graaf et al. 2006). However, a recent fMRI study revealed that the cerebellum is involved in the early phase of task performance and coordination since its activity diminishes as the task becomes well practiced (Hermsdorf et al. 2020). On the other hand, the functional role of the cerebellum has been mainly studied in motor adaptation tasks (Doppelmayr et al. 2016; Galea et al. 2011; Jayaram et al. 2012). Only one study investigated the causal role of the cerebellum in visuomotor sequence learning and showed that low-frequency rTMS over the lateral cerebellum resulted in a significant weakening in sequence learning (Torriero et al. 2004).
2. ‘Inhibitory’ and ‘Facilitatory’ rTMS Protocols
It is common to assume a linear relationship between the direction of the produced aftereffects on cortical excitability and the behavioral effects of rTMS. According to this view, low-frequency rTMS and cTBS (i.e., the ‘inhibitory’ protocols) might induce functional inhibition/disruption, whereas high-frequency rTMS and iTBS (i.e., the ‘excitatory’ protocols) might lead to functional improvements/enhancement. Although this generally accepted dichotomy between the stimulation frequency and the direction of the produced cognitive aftereffects is likely oversimplified, several studies discuss the results in this framework.
In the following, the researchers delineate several reasons that it is challenging to predict the functional aftereffects of rTMS solely based on the protocol type (i.e., ‘inhibitory’ or ‘excitatory’). First, there is substantial interindividual variability when inducing corticospinal excitability changes in M1. While group-level data might show frequency-dependent modulatory effects, they can vary significantly across individuals (Hamada et al. 2013; Maeda et al. 2000), and even within individuals (Goldsworthy et al. 2021). Biological (e.g., age, time of the day, genetics, brain state) and methodological factors (e.g., stimulation parameters, measures for the effect) both may be responsible for the intra- and interindividual variability of the rTMS effect (Huang et al. 2017). One possible strategy to decrease variability effects is increasing the specificity of the stimulation. This can be achieved by a novel rTMS technique, called quadripulse stimulation (QPS), that uses repetitive monophasic pulses, instead of biphasic pulses resulting in smaller variability in the after-effects (Simeoni et al. 2016; Tiksnadi et al. 2020).
Second, it is unclear whether a given protocol that may decrease the cortical excitability in M1 produces the same physiological effects in other cortical areas. For instance, some authors speculate that there might be overlaps in the produced aftereffects at least within the frontal cortex (e.g., M1 and DLPFC, as discussed in de Jesus et al. 2014). Third, this view may miss the brain’s endogenous and dynamic compensatory mechanisms to external perturbations. For example, due to the interhemispheric compensation, decreasing the excitability level of the left DLPFC with low-frequency rTMS may lead to the compensatory recruitment of the right DLPFC (Ambrus et al. 2020). Fourth, the stimulation frequency is only one of many crucial stimulation parameters that can shape the direction of aftereffects. For example, the facilitating effect of high-frequency rTMS requires inter-train intervals; otherwise, it is more likely to produce an inhibitory effect (Rothkegel et al. 2010).
Based on these arguments, it is conceivable to expect that the behavioral effects of rTMS may not always match the alterations in cortical excitability. Consequently, ‘facilitatory’ protocols may not always enhance, and ‘inhibitory’ protocols may not necessarily weaken the performance.
3. Stimulated Hemisphere(s)
Many studies have used rTMS to better understand the hemispheric involvement of a given brain region when performing the SRTT. These studies typically ask whether the left or right brain area (e.g., M1) is causally involved in a specific task phase (e.g., learning phase). To this aim, most studies have stimulated the left or the right hemisphere at a time and studied whether rTMS could modulate the performance.
Performing the SRTT may require using only one hand or both hands. When the participants perform the SRTT with only one hand (e.g., the right hand), the stimulation may target the contralateral (i.e., left) or ipsilateral (i.e., right) hemisphere. Eleven of the 17 studies targeted the contralateral hemisphere (see Table 1). In studies where only the left hemisphere was stimulated, only right-handed participants were included (see Table 1).
Table 1. The effect of different rTMS protocols on visuomotor sequence learning according to the target area, sequence type, and timing of the stimulation.
| Target Area | Authors and Year | rTMS Protocol | Hemisphere | Timing of Stimulation | Type of Sequence | Outcome |
|---|---|---|---|---|---|---|
| M1 | ||||||
| (Robertson et al. 2005) | 1 Hz rTMS | Left only | After learning | 12-item implicit deterministic | Blocked offline improvements over the day, but not overnight | |
| (Breton and Robertson 2017) | 1 Hz rTMS | Left only | After learning | 12-item implicit/explicit deterministic | Blocked offline improvements in explicit, but not in implicit task | |
| (Tunovic et al. 2014) (Experiment 3) | cTBS | Right only | After learning | 12-item explicit deterministic | Offline improvements after cTBS | |
| (Cohen and Robertson 2011) (Experiment 2) |
1 Hz rTMS | Right only | After learning | 12-item implicit deterministic | Increased learning after 12 h consolidation by preventing interference with a declarative task | |
| (Wilkinson et al. 2010) | cTBS, iTBS | Left only | Before learning | 12-item implicit probabilistic | Learning was prevented by cTBS | |
| (Wilkinson et al. 2015) | cTBS | Left only | Before learning | 12-item implicit probabilistic | Decreased initial sequence learning and recall | |
| (Steel et al. 2016) | cTBS | Left only | Before learning | 12-item implicit probabilistic | Learning was disrupted | |
| (Rosenthal et al. 2009) (Experiment 1, 2) |
cTBS | Contralateral to dominant hand | Before learning | 12-item implicit probabilistic | Learning was disrupted in manual, but not in perceptual task | |
| (Clark et al. 2019) | cTBS | Left only | Before learning | 12-item implicit deterministic | Decreased learning in simple, but not in a more complex sequence | |
| DLPFC | ||||||
| (Pascual-Leone et al. 1996) | 5 Hz rTMS | Left or right in separate conditions | During learning | 12-item implicit deterministic | Learning was disrupted | |
| (Robertson et al. 2001) | 1 Hz rTMS | Contralateral to dominant hand | Before learning | 10-item implicit deterministic | Learning was prevented in spatial, but not in color cue guided task | |
| (Wilkinson et al. 2010) | cTBS | Left only | Before learning | 12-item implicit probabilistic | No effect on learning | |
| (Gann et al. 2021) | cTBS, iTBS | Left only | Before learning | 8-item explicit deterministic | No effect on learning | |
| (Galea et al. 2010) | cTBS | Left or right in separate groups | After learning | 12-item explicit deterministic | Improved learning after 8 h consolidation | |
| (Tunovic et al. 2014) (Experiment 2) |
cTBS | Right only | After learning | 12-item explicit deterministic | Offline improvements after cTBS | |
| (Cohen and Robertson 2011) (Experiment 2) |
1 Hz rTMS | Right only | After learning | 12-item implicit deterministic | Decreased learning after 12 h consolidation by failing to prevent interference with a declarative task | |
| (Ambrus et al. 2020) | 1 Hz rTMS | Bilaterally | Between learning blocks | 8-item implicit probabilistic | Improved learning after 24 h consolidation | |
| Broca’s area | ||||||
| (Clerget et al. 2012) | cTBS | Left only | Before learning | 20-item implicit deterministic | Learning was prevented | |
| SMA | ||||||
| (Pascual-Leone et al. 1996) | 5 Hz rTMS | Not applicable | During learning | 12-item implicit deterministic | No effect on learning | |
| (Wilkinson et al. 2010) | cTBS | Not applicable | Before learning | 12-item implicit probabilistic | No effect on learning | |
| (Perez et al. 2008) | 1 Hz rTMS | Not applicable | During learning | 12-item implicit deterministic | Blocked intermanual transfer of the skill | |
| IPL | ||||||
| (Robertson et al. 2001) | 1 Hz rTMS | Contralateral to the dominant hand | Before learning | 10-item implicit deterministic | No effect on learning | |
| (Rosenthal et al. 2009) (Experiment 1, 2) |
cTBS | Right only | Before learning | 12-item implicit probabilistic | Learning was disrupted in perceptual, but not in manual task | |
| (Breton and Robertson 2017) | 1 Hz rTMS | Left only | After learning | 12-item implicit, explicit deterministic | Blocked offline improvements in implicit, but not in explicit task | |
| (Clark et al. 2019) | cTBS | Left only | Before learning | 12-item implicit, deterministic | No effect on learning | |
| Cerebellum | ||||||
| (Torriero et al. 2004) | 1 Hz rTMS | Left or right in separate groups | Before learning | 12-item implicit, deterministic | Learning was disrupted |
However, studies targeting the dominant hemisphere (based on M1) may neglect the possibility that the non-stimulated hemisphere can take over the function of the stimulated one (Andoh and Martinot 2008; Sack et al. 2005), potentially influencing the results. Applying sequential bilateral stimulations (i.e., delivering the same rTMS protocol over a given cortical target consecutively on each hemisphere) may be a promising solution to overcome the possible interhemispheric compensatory mechanisms (Ambrus et al. 2020).
Considering the side of the stimulation, some studies have targeted both hemispheres in separate experimental groups. Using this method, Galea et al. (2010) successfully demonstrated that cTBS over the right DLPFC improved visuomotor sequence learning to a greater extent than the left DLPFC (Galea et al. 2010). Another study revealed a dissociation between cerebellar hemispheres: the stimulation of the right cerebellar hemisphere weakened sequence learning regardless of which hand was used, while the interference with the left cerebellar hemisphere affected only through the ipsilateral hand (Torriero et al. 2004). Therefore, this method is suitable for exploring potential lateralization effects as well.
For non-motor brain areas, it is worth targeting both hemispheres separately and applying sequential bilateral protocols. This approach can avoid hemispheric compensatory mechanisms and reveal the possible dissociation between hemispheres (Ambrus et al. 2020). Furthermore, the sequential bilateral stimulation for the two-handed version of the task may be a particularly good solution.
4. Timing of the Stimulation
Motor memory traces can be strengthened in two ways: when performance is gradually improving during practice (online learning) and when performance improves between two training sessions without any practice (offline learning) (Cohen et al. 2005; Robertson et al. 2004). Sequence learning is a multi-stage process: it consists of the learning, consolidation, and retrieval phases.
Learning phase. One can deliver rTMS immediately before or during the learning phase (see Figure 4). During the learning phase, rTMS may be applied simultaneously with the task performance or between the learning blocks. Only two studies have applied rTMS during the initial sequence learning process (so-called ‘online stimulation’). In one study, high-frequency rTMS over the DLPFC led to a performance decrease on the SRTT (Pascual-Leone et al. 1996). In a more recent study, the authors applied low-frequency rTMS over the DLPFC between the learning blocks and found a performance improvement on an alternating SRTT (Ambrus et al. 2020). Most research delivered stimulation immediately before task performance (so-called ‘offline stimulation’; see Table 1). Most of them have found decreased learning of visuomotor sequence learning (Clark et al. 2019; Clerget et al. 2012; Torriero et al. 2004; Wilkinson et al. 2015).
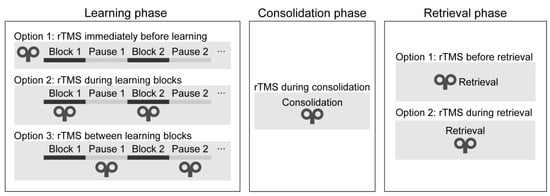
Figure 4. Timing of the stimulation in each task phase.
Consolidation phase. One may apply rTMS after the learning phase to verify its effect on memory consolidation (see Figure 4). In one study, cTBS over the DLPFC improved performance after an 8-h-long offline period (Galea et al. 2010). In contrast, low-frequency rTMS over M1 blocked offline improvements on an implicit SRTT over the day (Robertson et al. 2005), as well as on an explicit SRTT after sleep (Breton and Robertson 2017). rTMS over the DLPFC and M1 can influence the development of memory traces because their stimulation leads to changes in the consolidation process.
Retrieval phase. Finally, rTMS may be applied immediately before or during the recall phase. No studies have applied rTMS immediately before or during the recall phase as of the day of the literature search. Thus, it is unclear whether rTMS can modulate the recall of well-acquired sequence knowledge.
5. Type of the SRTT Sequence
The effect of stimulation may depend on the SRTT sequence type. Several new versions of the SRTT have emerged that can differ in three crucial dimensions: the sequence applied can be (1) implicit or explicit, (2) deterministic or probabilistic, and (3) first-order conditional (FOC) or second-order conditional (SOC) (as defined in section Different variations of the SRTT).
Implicit vs. explicit sequences. The most commonly used version of the SRTT uses implicit sequences (here, 13 out of the 17 reviewed studies used an implicit SRTT). However, its explicit version (i.e., the existence of the predetermined sequence is revealed to the participants before learning) can also be applied if the goal is to test intentional learning or declarative knowledge of the sequence. They examined the role of M1 and the IPL in both the implicit and explicit SRTT and revealed a double dissociation: low-frequency rTMS over the IPL prevented offline improvement in the implicit but not in the explicit task. On the other hand, the same stimulation over M1 prevented offline improvement in the explicit but not in the implicit task (Breton and Robertson 2017).
Deterministic vs. probabilistic sequences. In the classic SRTT, stimuli follow a fixed order, creating a deterministic sequence (Nissen and Bullemer 1987; Shanks 2005). Probabilistic types of the SRTT also exist, where the sequence is hidden in noise; therefore, learning is more likely to remain implicit (Howard et al. 2004; Song et al. 2007). Five out of 17 studies used probabilistic sequences, and 12 used deterministic sequences (Table 1). All of the studies employing probabilistic sequence learning tasks found behavioral effects of rTMS. In four studies, learning deteriorated (Rosenthal et al. 2009; Steel et al. 2016; Wilkinson et al. 2010, 2015), and, in one study, learning was improved (Ambrus et al. 2020). On the other hand, deterministic sequence learning performance was successfully manipulated in 11 out of the 12 studies: in eight studies, learning was disrupted (Breton and Robertson 2017; Clark et al. 2019; Clerget et al. 2012; Pascual-Leone et al. 1996; Perez et al. 2008; Robertson et al. 2005, 2001; Torriero et al. 2004), and, in three studies, learning was improved (Cohen and Robertson 2011; Galea et al. 2010; Tunovic et al. 2014). Therefore, it seems that rTMS can equally modify the learning of both probabilistic and deterministic sequences.
FOC vs. SOC sequences. Another critical factor is the statistical structure of the sequence. In the simpler first-order conditional (FOC) sequences, elements can be predicted by the preceding one. On the other hand, in the more complex second-order conditional (SOC) sequences, it is the combination of two consecutive elements that predicts the forthcoming one. Clark et al. (2019) investigated the role of M1 in the acquisition of simpler FOC and more complex SOC sequences. According to their findings, cTBS over M1 resulted in poorer learning of the FOC sequence compared to the SOC sequence. These findings support the hypothesis that the acquisition of FOC and SOC sequences may rely on different neural networks: simpler FOC sequences are processed by a circuitry involving M1, while more complex SOC sequences are associated with an expanded network, including Brodmann area 44 (BA44) and the DLPFC (Ashe et al. 2006; Lum et al. 2018). Although working with discrete sequence production task instead of the SRTT, an rTMS study also revealed the distinct role of the pre-SMA in more complex sequences compared to simpler ones (Ruitenberg et al. 2014). Based on these promising results, future studies may investigate FOC and SOC sequences targeting non-motor areas too.
This entry is adapted from the peer-reviewed paper 10.3390/jintelligence11100201
This entry is offline, you can click here to edit this entry!