1. Introduction
Cancer cells are discussed to be “addicted” to the sulfur-containing essentiell amino acid methionine [7]. The selective upregulation of the methionine transporter SLC43A2 allows cancer cells to outcompete cells in the tumor microenvironment for methionine supply [8]. Methionine is used for the biosynthesis of proteins, nucleotides, glutathione, and the methyl donor S-adenosylmethionine (SAM), which are required for cellular growth and proliferation, as well as protection against oxidative stress [7]. Moreover, methionine serves as a precursor for the endogenous and exogenous generation of two volatile sulfur compounds (VSCs): hydrogen sulfide (H2S) and methanethiol. Several strains of gut bacteria may convert dietary methionine to H2S and/or methanethiol, thus representing the major source of exposure of humans to both VSCs. In addition, H2S and methanethiol are produced endogenously through enzymatic and non-enzymatic synthesis [9,10]. The cancer-associated dysregulation of sulfur metabolism results in excess levels of both VSCs in tumor tissue as well as in body fluids, breath, and/or excretions of cancer patients that are increasingly being exploited for the establishment of convenient non-invasive screening methods to detect early signs of cancer [11–14]. In this regard, the dysregulated H2S biosynthesis in various types of cancer and the cancer-promoting effects of elevated H2S levels have recently been discussed in several reviews [10,15,16]. Excess H2S may promote cellular dedifferentiation and the growth of transformed cells as well as increase their potential to metastasize and develop resistance against chemotherapeutic agents [10]. The endogenous production of H2S in mammalian cells occurs largely through four enzymes: cystathionine β-synthase (CBS), cystathionine γ-lyase (CTH), 3-mercaptopyruvate sulfurtransferase (MPST), and selenium-binding protein 1 (SELENBP1) [9,17,18]. In particular, CBS has been shown to be upregulated in various types of cancer, including colorectal carcinoma (CRC), squamous cell carcinoma, and ovarian, breast, and thyroid cancers [10]. In several of those cancers, CTH and MPST levels were also found to be elevated [10]. On the other hand, SELENBP1, which generates H2S through the oxidation of methanethiol [18], is often downregulated in tumor tissue [19].
Methanethiol (methyl mercaptan, CH3SH) is an alkyl thiol and a member of a group of repulsive-smelling VSCs, characterized by their toxicity and low odor threshold. It is a colorless gas with a boiling point of 5.9 °C. The smell of methanethiol is reminiscent of rotten cabbage and is perceptible to humans at concentrations of ~1–2 parts per billion (ppb) [20]. A study on rats that investigated the toxicity of different VSCs reported a 24 h LD50 value of 675 parts per million (ppm) for acute exposure to methanethiol, which was similar to the LD50 of 444 ppm measured for H2S [21]. The toxicity of methanethiol has been attributed to its inhibiting effects on cytochrome c oxidase and the electron transfer in the respiratory chain [22]. Environmental microbial production of methanethiol occurs mainly through the methylation of H2S in the anoxic sediment/water interphase and through the elimination reactions of sulfur-containing amino acids catalyzed by L-methionine-γ-lyase (MGL). In addition, it can be generated from dimethylsulfoniopropionate (DMSP), an osmolyte in marine algae, through a coupled series of bacterial demethylation and cleavage reactions [20]. Although methanethiol has long been known as an environmental toxin and an intermediate in the biogeochemical sulfur cycle, its metabolism in humans has only recently gained more attention, following the identification of human selenium-binding protein 1 (SELENBP1) as a novel methanethiol-oxidizing enzyme in 2018 [18].
2. Exogenous and Endogenous Methanethiol Production in Humans
Methanethiol is a common volatile component of the human flatus: in healthy volunteers to whom beans and lactulose were given to enhance their flatus output, the measured methanethiol concentrations reached 0.19–0.24 µmol/L (9.14–11.55 ppb). Thus, among the detected VSCs, methanethiol showed the second highest concentrations after H
2S [
23]. Analogous to H
2S, the majority of methanethiol biosynthesis in humans likely originates from the gut microbiome. Commensal bacteria residing in the colonic lumen can convert dietary methionine to methanethiol through MGL-catalyzed α,γ-elimination and γ-replacement reactions.
Fusobacterium nucleatum,
Citrobacter freundii,
Morganella morganii, and several
Proteus species have been reported to contribute to intestinal methanethiol production [
24,
25]. Individual variations in the composition of the bacterial strains in the colon as well as differences between human populations may, thus, affect the exogenous production of methanethiol. The major role of gut microbiota as methanethiol producers is supported by the demonstration of decreased methanethiol levels in patients who were treated with antibiotics [
18]. Methanethiol concentrations released by the intestinal flora are dependent on nutritional factors and may be influenced by the type of diet and dietary restrictions [
23]. In particular, the ingestion of dietary proteins that contain high amounts of methionine may result in increased methanethiol production by the gut microbiome, whereas activated charcoal and zinc salts have been demonstrated to neutralize sulfur-containing malodorous gases such as methanethiol and H
2S [
23]. Besides being excreted through flatus and feces, some of the bacteria-derived methanethiol diffuses into the epithelial cells lining the colonic lumen of the human host [
9].
Regarding endogenous production, there is experimental evidence for the occurrence of two methanethiol-generating enzymatic pathways in human cells. Recently, recombinant methyltransferase-like protein 7B (METTL7B;
aka thiol S-methyltransferase TMT1B) has been demonstrated to be capable of producing methanethiol through the methylation of H
2S, using SAM as the methyl donor [
26]. METTL7B is highly abundant in the gut, the liver, the kidneys, and the lungs (
https://www.proteinatlas.org/ENSG00000170439-METTL7B/tissue, accessed on 3 July 2023); nevertheless, the quantitative contribution and the physiological relevance of this novel methanethiol-generating enzymatic pathway remain to be determined. Furthermore, a study on isolated mitochondria found that methanethiol may arise from the oxidation of the transamination product of methionine, α-keto-methylthiobutyrate [
27]. However, this pathway might be quantitatively relevant only under conditions of methionine excess. In this regard, mice fed a methionine-rich diet showed characteristic features of methanethiol toxicity [
28]. Polymorphisms in the genes, coding for the enzymes that are responsible for endogenous methanethiol production, may also have an influence on the cellular and systemic methanethiol concentrations; however, this remains to be explored yet.
3. SELENBP1-Catalyzed Degradation of Methanethiol in Humans
As mentioned above, exposure of humans to methanethiol is thought to occur mainly via the distal gastrointestinal tract due to the degradation of dietary methionine by gut microbiota. The rapid detoxification of methanethiol in the colonic mucosa through oxidative conversion to H
2S and, further on, to thiosulfate has already been reported, more than 20 years ago [
29,
30], but the executing enzymes remained elusive at that time. In 2018, SELENBP1 was identified to act as a methanethiol oxidase (MTO), catalyzing the oxidation of methanethiol to formaldehyde, hydrogen peroxide (H
2O
2), and H
2S in the presence of oxygen (
Figure 1) [
18]. Subsequently, H
2S is metabolized to thiosulfate and sulfate in the sulfide oxidation unit, comprising four enzymes located in mitochondria [
31].
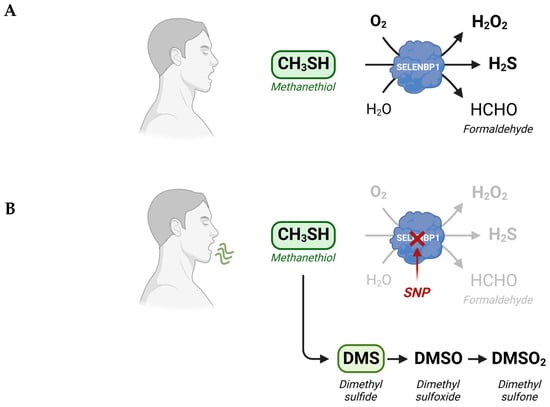
Figure 1. Alternate metabolic fates of methanethiol. (
A) Physiologically, degradation of methanethiol occurs predominantly through SELENBP1, which acts as MTO, catalyzing the rapid oxidative conversion of methanethiol to H
2S, H
2O
2, and formaldehyde. (
B) In SELENBP1-deficient cancer cells and in cells that possess an inactive MTO due to mutations (SNPs) in the
SELENBP1 gene, methanethiol is methylated to DMS, which can be oxidized subsequently to DMSO and DMSO
2. Methanethiol and DMS may be detected in the exhaled breath of afflicted persons. Scheme created with
Biorender.com.
Even though SELENBP1 is ubiquitously expressed in human tissues, it is particularly abundant in the intestinal epithelium (
https://www.proteinatlas.org/ENSG00000143416-SELENBP1/tissue, accessed on 5 July 2023), with a gradient in SELENBP1 expression along the crypt-luminal axis [
38]. Immunohistochemical analysis of human colon biopsies from healthy donors revealed that SELENBP1 levels were highest in the epithelial cells located at the tip of the villi [
38], which are in close contact with substances entering from the colonic lumen. Besides microbiota-derived methanethiol, SELENBP1 may also accept structurally related alkyl thiols as substrates, some of which arise from the digestion of dietary sulfur-containing phytochemicals [
32].
Cellular differentiation appears to be a major trigger of SELENBP1 expression: both the spontaneous and butyrate-induced differentiation of proliferating Caco-2 cells, a human intestinal adenocarcinoma cell line, to a colonocyte-like phenotype is associated with induction of SELENBP1 gene and protein expression and an increase in MTO activity in the terminally differentiated cells [
9,
38,
39]. The downregulation of SELENBP1 via the treatment of Caco-2 cells with small interfering RNA (siRNA) resulted in lowered expression of a differentiation marker of colonic epithelial cells, carcinoembryonic antigen [
38]. SELENBP1 also became upregulated in the course of differentiation of HT29 cl.16E cells, an in vitro model of intestinal secretory cells, to a goblet-cell-like phenotype [
38]. In vivo, an increase in SELENBP1 levels has been observed to occur along the colonic crypt–luminal axis, with the terminally differentiated epithelial cells at the top of the crypts showing the highest SELENBP1 expression [
38]. Moreover, SELENBP1 has been reported to be a marker protein of terminally differentiated erythrocytes and adipocytes [
40,
41]. SELENBP1 is a highly abundant non-heme protein in erythrocytes [
42], where it is presumably responsible for the degradation of circulating methanethiol. Indeed, MTO activity in whole blood has been attributed predominantly to the erythrocyte fraction [
18]. Regarding adipocytes, SELENBP1 appears to be not only a marker but also an endogenous stimulator of terminal differentiation that is promoted by the signaling mediator H
2S: the knockdown of SELENBP1 in murine 3T3-L1 pre-adipocytes, thus, resulted in the suppression of common features of adipocyte differentiation, such as intracellular lipid accumulation and the induction of adiponectin expression [
43].
Two of the products of the SELENBP1-catalyzed oxidation of methanethiol, H
2O
2, and H
2S, may exert pleiotropic effects and bivalent (both stimulatory and inhibitory) actions on signaling pathways, depending on their concentration and cellular localization as well as the (patho)physiological context. Both molecules are cytotoxic at high concentrations, whereas they are implicated as second messengers in redox signaling at lower concentrations. Thus, they may affect a wide variety of cellular processes, including differentiation, apoptosis, antioxidant adaptation, and mitochondrial respiration [
44,
45]. In addition, H
2S may serve as an electron donor for the generation of ATP in the mitochondrial respiratory chain [
46]. However, it still needs to be explored to what extent SELENBP1 contributes to the generation of H
2O
2 and H
2S in different types of cells and whether biological effects of SELENBP1 other than methanethiol detoxification could be attributed to the MTO reaction products.
In addition to the intestine, several other tissues in the human body show high SELENBP1 levels, including the liver, the lungs, and the nasopharynx (
https://www.proteinatlas.org/ENSG00000143416-SELENBP1/tissue, accessed on 5 July 2023). The liver appears to be involved in the detoxification of circulating methanethiol, as elevated concentrations of methanethiol were measured in the blood of patients suffering from liver cirrhosis with and without hepatic encephalopathy, and increasing methanethiol concentrations over time were associated with a progressive decline in the health status of the patients [
47]. Also, the lungs might be capable of metabolizing methanethiol circulating in the blood, while methanethiol produced by bacteria populating the oral cavity might be detoxified in the mucosa of the nasopharynx. In this regard, elevated methanethiol concentrations have been measured in the oral cavity of patients with chronic periodontitis and intra-oral halitosis [
48].
Interestingly, SELENBP1 was identified to act as MTO in a study based on five patients suffering from extra-oral halitosis, who were found to possess biallelic single-nucleotide polymorphisms (SNPs) in the
SELENBP1 gene. The patients showed up with cabbage-like malodor, due to high levels of methanethiol and another VSC, dimethyl sulfide (DMS), in their breath. The detected point mutations caused exchanges of single amino acids (Gly225Trp, His329Tyr) in SELENBP1 that resulted in a loss of MTO activity and, in turn, the accumulation of methanethiol and DMS in breath and body fluids. DMS is generated via the methylation of methanethiol under MTO-deficient conditions (
Figure 1). The homologous overexpression of wild-type SELENBP1 restored the MTO activity in fibroblasts prepared from one of the patients [
18].
4. Elevated Levels of Methanethiol and Its Derivatives in Various Types of Cancer
Both the generation and metabolism of methanethiol can be altered/dysregulated in cancer, resulting in elevated levels of methanethiol and methanethiol-derived metabolites in tumor tissue. Since methanethiol is highly volatile and membrane-permeant, the footprint of its dysregulated metabolism is found in both the body fluids, such as blood and urine, and excreted gases, such as breath and flatus, of cancer patients.
Differences in gut microbiota with respect to composition and relative abundance have been reported between healthy persons and patients with CRC [
49]. Among the methanethiol-generating bacteria in the colon,
Fusobacterium nucleatum was over-represented in CRC patients [
50]. Correspondingly, higher methanethiol concentrations have been measured in the flatus of CRC patients, as compared to healthy individuals [
13,
51].
Elevated methanethiol concentrations were also found in the exhaled breath of persons diagnosed with oral squamous cell carcinoma [
52], probably deriving from the high endogenous synthesis in the tumor tissue. The uptake of glucose and methionine is often elevated in cancer cells due to the upregulation of the respective transporters [
5,
6,
7]. The accumulation of these two metabolites in cancer cells may favor a Maillard reaction between them, resulting in the non-enzymatic synthesis of methanethiol and its subsequent release, as demonstrated in in vitro experiments [
51]. Moreover, methionine serves as a precursor for the biosynthesis of the methyl donor SAM that is catalyzed by the enzyme methionine adenosyl transferase 2A (MAT2A). Elevated MAT2A levels were observed in various types of cancer, including CRC, hepatocellular carcinoma (HCC), and breast and endometrial cancers, and are mostly considered as an unfavorable prognostic marker (
https://www.proteinatlas.org/ENSG00000168906-MAT2A/pathology, accessed on 17 July 2023) [
53]. As mentioned above, METTL7B may use SAM to produce methanethiol via the methylation of H
2S [
26]. Like MAT2A and the methionine transporter SLC43A2, METTL7B is upregulated in several types of cancer (
https://www.proteinatlas.org/ENSG00000170439-METTL7B/pathology, accessed on 17 July 2023), thus providing a molecular rationale for elevated methionine-derived enzymatic methanethiol production in cancer cells. In thyroid cancer, where METTL7B is highly expressed, METTL7B has recently been shown to promote metastasis by increasing migration and invasion [
54].
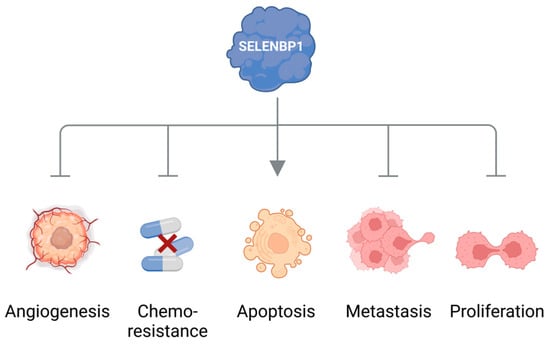
Another strong hint pointing to the dysregulation of methanethiol metabolism in cancer stems from observations that the methanethiol-degrading enzyme SELENBP1 is often and markedly downregulated in cancer tissue. Lowered expression of SELENBP1 has been reported for many types of cancer, such as tumors of the colon, lungs, ovaries, prostate, liver, thyroid, kidneys, and breast [
19]. SELENBP1 has been designated a tumor suppressor that may inhibit cell proliferation, angiogenesis, metastasis, and resistance to chemotherapy, as well as promote apoptotic cell death (
Figure 2) [
19,
55,
56,
57,
58,
59]. Moreover, low SELENBP1 levels in cancer tissue correlate with poor clinical prognosis for the patients [
19].
Figure 2. SELENBP1 as a tumor suppressor. SELENBP1 has been reported to inhibit cell proliferation, angiogenesis, metastasis, and resistance to chemotherapy as well as to promote apoptotic cell death. Scheme created with
Biorender.com.
MTO activities in tumor tissues of patients have not been assessed yet; however, severely decreased levels of an enzyme are usually associated with a diminished capacity to convert its substrate. In this regard, the erythrocytes and different tissues of SELENBP1-knockout (KO) mice were shown to be deficient in MTO activity and DMS, a biomarker of defective methanethiol oxidation accumulated in the plasma of the KO mice. Compared to the wild-type mice, even the heterozygous carriers exhibited less MTO activity in their erythrocytes and moderately elevated plasma levels of DMS [
18]. It can be assumed that the catabolism of methanethiol in cancer patients is similarly shifted from predominant oxidation to methylation. Under the conditions of low SELENBP1 and high METTL7B expression in cancer cells, METTL7B is then likely to catalyze the methylation of a part of the excessive methanethiol to DMS. Indeed, DMS levels were reported to be elevated in the breath of patients with HCC and lung cancer, as compared to healthy controls [
60,
61]. High concentrations of DMS were also measured in the headspace of the tumor-derived human HepG2 hepatoma and A549 lung adenocarcinoma cell lines [
62]. Some DMS may be oxidized to dimethyl sulfoxide (DMSO) and, further on, to dimethyl sulfone (DMSO
2) (
Figure 1) [
18]. Elevated levels of DMSO
2, the stable end product of this pathway, were detected in melanomas and endometrial and ovarian carcinomas [
63,
64,
65]. Polysulfides, such as dimethyl trisulfide, are additional products of a dysregulated methanethiol metabolism and have been found among the volatiles emitted by melanoma cells [
63]. Following subcutaneous injection of melanoma cells, both DMSO
2 and dimethyl trisulfide concentrations were found to be increased in the urine of tumor-bearing mice, as compared to healthy controls [
66].
Besides being excreted or methylated, excessive methanethiol may modify cysteine residues in proteins, as demonstrated for a methanethiol adduct of Cys34 in human albumin, which was more abundant in the serum of CRC patients compared to healthy persons [
67].
Cancerous ulcers are probably the most suitable tumor tissue for the direct detection of characteristic volatiles in cancer emissions since their volatilome is not metabolized prior to release. Cancerous ulcers, or fungating cancer, occur when malignant tumor cells invade and erode the skin. This is often associated with the emission of unpleasant odors. The predominant odor-producing component in fungating head, neck, and breast cancers has been identified as the methanethiol-derived metabolite dimethyl trisulfide [
68,
69]. In this regard, it should be noted as well that ulcers are often infected, and a microbial origin of the detected dimethyl trisulfide cannot be ruled out.
5. Conclusions and Outlook: Methanethiol as a Promising Biomarker for Non-Invasive Cancer Diagnosis
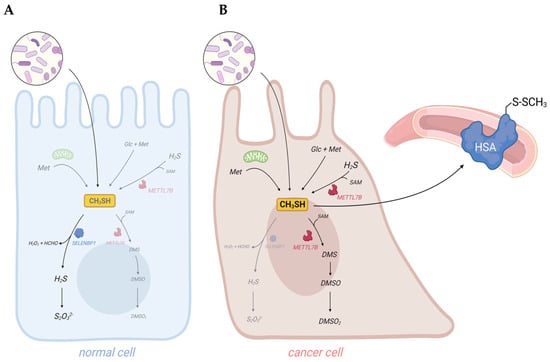
Dysregulated metabolism in cancer cells results in the generation of cancer-associated volatile organic compounds (VOCs). Their measurement in body fluids, exhaled breath, and flatus may provide a tool for early non-invasive diagnosis of cancer. Among the identified cancer-associated VOCs are hydrocarbons, alcohols, aldehydes, ketones, and VSCs, such as H2S and methanethiol [11–13,70–72]. Indeed, artificial intelligence (AI)-based surface-enhanced Raman spectroscopy (SERS) has recently been demonstrated as a useful and accurate method for the early diagnosis of oral cancer through the detection of methanethiol in exhaled breath [14]. Elevated concentrations of methanethiol in cancer patients may derive from increased exogenous and endogenous synthesis and suppressed degradation. Moreover, the downregulation of the methanethiol-oxidizing enzyme SELENBP1 that is observed in many types of cancer may shift its metabolism to the generation of methylated derivatives, which have been detected in cancer patients as well (Figure 3).
Taken together, elevated concentrations of methanethiol and its methylated derivatives in body fluids and excreted gases contribute to the “scent of cancer”, frequently observed in affected patients.
Figure 3. Dysregulation of sulfur metabolism in cancer cells may result in elevated levels of methanethiol and its derivatives in body fluids and excreted gases of cancer patients. (
A) In healthy persons, the vast majority of methanethiol derives from the metabolic activity of gut bacteria. Some methanethiol may diffuse into the colonic epithelium, where it is rapidly oxidized by SELENBP1. (
B) In cancer patients, endogenous synthesis of methanethiol may substantially increase due to elevated levels of metabolites, such as methionine (Met), glucose (Glc), H
2S, and SAM, in tumor cells. Downregulation of SELENBP1 and upregulation of METTL7B may result in a shift from oxidation to methylation of methanethiol. As methanethiol and its methylated derivatives are highly volatile, they may be used as biomarkers for non-invasive cancer detection. Adducts of methanethiol with proteins may also be detected, as it has been demonstrated for human serum albumin (HSA). Scheme created with
Biorender.com.
This entry is adapted from the peer-reviewed paper 10.3390/antiox12091780