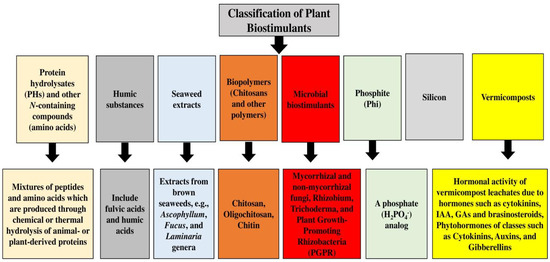
Biostimulants are classified into two distinct groups based on their origin; one category includes products that have biological origins in pathogens or plants, and the second group consists of products that do not have biological origins [
36,
37,
38]. Another classification approach divides biostimulant products into microbial biostimulants, which are obtained from arbuscular mycorrhizal fungi and plant-growth-promoting bacteria, and non-microbial biostimulants, which include plant micro-algae extracts, humic substances, and biopolymers such as chitosan [
39,
40,
41,
42,
43]. Different compounds with bioactive properties can be used as biostimulants to boost plant growth and development under normal and stress conditions [
44,
45,
46,
47,
48,
49,
50,
51,
52]. Salicylic acid is economical and quick in action, environmentally sound, and it also links with other elicitors to boost the biosynthesis of secondary metabolites [
53,
54,
55]. Humic acid can increase plant growth, retain water, enrich nutrients, and suppress disease [
56,
57]. Fulvic acids are used in sustainable horticulture and can change plant primary and secondary metabolism and increase nutrient uptake, root growth, and crop tolerance to environmental stresses [
58,
59]. Protein hydrolysate biostimulants, mostly produced by chemical and enzymatic hydrolysis of plant- and animal-derived proteins, are based on a mixture of soluble amino acids and peptides and can increase the yield and quality of products as well as improve the nutrient uptake and abiotic stress tolerance of plants [
60,
61,
62,
63]. They are largely prepared from brown seaweeds, such as
Ecklonia maxima,
Ascophyllum nodosum, and
Macrocystis pyrifera, and they include promoting hormones or trace elements such as Zn, Fe, Mn, and Cu [
64,
65]. Humic-like substances such as fulvic and humic acids may also show biostimulatory activity, since several reports have suggested improved crop performance attributed mainly to auxin- and cytokinin-like impacts; they are obtained from organic matter decomposition and metabolic products of soil microbes, and they have roles in plant growth via the improvement of soil physical–chemical properties and the boosted availability of nutrients in the rhizosphere [
66,
67,
68]. The actual mechanisms of action seem to be the result of synergy between the several bioactive components in raw materials, although the impacts may change depending on the crop, soil type, and soil microbes present in the rhizosphere [
69,
70,
71,
72]. The most important impacts of chitin and its derivatives’ applications are that they stimulate and protect seed germination, stimulate stress resistance, mitigate negative impacts of abiotic stress, induce plant growth and development, improve soil properties and prevent nutrient leaching, improve the shelf-life of crops, chelate heavy metals, increase crop yield and quality, and protect against pests and pathogens, e.g., bacteria, viruses, fungi, insects, and nematodes [
73,
74,
75]. Amino acids are the best candidates to boost stress tolerance through osmo-protection, ROS scavenging, metal chelation, and nutrient availability [
76], which can notably impact the synthesis and stimulation of some enzymes and gene expression [
77,
78,
79]. They can also be applied as signal molecules, like for inducing stomatal closure, as sensors of the nutrient contents of cells, or as regulators for inducing their own catabolism. Amino acids can manage the procedure of protein synthesis, strengthening plant growth, photosynthesis, and yield formation. They can increase nutrient assimilation, use, and translocation, as well as increase the quality of constituents [
79]. Amino acids are well-known biostimulants due to their positive effects on yield and plant growth, and can mitigate injuries from abiotic stresses [
80,
81]. Amino acids also have a significant role in ammonium fixation and C
4 metabolism and in the biosynthesis of different components, including isoflavonoids, flavonoids, cutin, aurones, sporopollenin, stilbenes, proanthocyanidins, suberin, lignins, catechins, phenylpropenes, lignans, acylated polyamines, and other different alkaloid derivatives. The largest and most diverse group of secondary metabolites in plants is phenols, which have good antioxidant effects and are involved in the regulation of photosynthesis, physiological activities, oxidation reduction procedures, and plant breathing [
82]. Phenolic acids and their derivatives are coumarins, stilbenes, quinones, lignans, flavonoids, curcuminoids, and tannins, which have meaningful roles in plant development, especially in pigment and lignin biosynthesis, and of course, they have a significant role in protecting plants from stress [
83].
Protein hydrolysate biostimulants, mostly produced by enzymatic and chemical hydrolysis of plant-derived and animal proteins, are based on a mixture of peptides and soluble amino acids, and can increase the quality and yield of products as well as the uptake and abiotic stress tolerance of plants [
84].
Glomus, the largest and most common genus in the phylum
Glomeromycota, forms symbiotic relationships with plant roots [
85,
86], which can boost the drought tolerance of the host plant, mediated by proteins with chaperone-like activity [
87].
Trichoderma fungi have important functions in nature as plant growth promoters and antagonists of phytopathogenic fungi [
88], and as rhizosphere inhabitants, they contribute to interactions with microorganisms, soil, arthropods, and plants at multiple trophic levels [
89], and can be used as biocontrol and biopesticide agents [
90]. Members of the genus
Trichoderma are also used in different industry branches, like in the production of biofuel, antibiotics, and enzymes [
91]. The main
Trichoderma–plant interactions include their impacts on plant morphology, plant physiology, nutrient absorption and solubilization, disease resistance, yield improvement, and abiotic stress tolerance [
92].
Trichoderma reesei is a genus of filamentous fungi and a superior cellulose source for industrial uses, and it can produce proteins, including different enzymes, cellulases, hemicellulases, and hydrophobins [
93,
94]. The endophytic fungus
Heteroconium chaetospira can also penetrate through the outer epidermal cells of its host, pass into the inner cortex, and grow all over the cortical cells, consisting of those of the root tip region, without causing apparent pathogenic symptoms [
95], and it can provide even more nitrogen to the plant than mineralizing plant-available organic nitrogen [
96].
Arthrobacter species, which are Gram-positive chemoorganotrophs and obligate aerobes, are commonly identified among soil bacteria [
97], being dominant aerobic bacteria under the class of families
Micrococcaceae and
Actinobacteria [
98], and nutritional versatility is the principal feature of arthrobacters [
99].
Acinetobacter spp. are Gram-negative coccobacilli that are aerobic, non-motile, and oxidative negative, with no glucose fermentation ability; they can be found in different environments [
100] and can fix nitrogen, solubilize minerals, produce siderophores, and act as plant endophytes or epiphytes, which can help hosts in detaching pollutants and tolerating environmental stresses [
101]. Moreover, the plant-growth-promoting traits of
Actinobacteria entail phosphate solubilization, IAA, and siderophores [
102]. They can also promote higher phosphorus content and plant growth and increase radical scavenging, plant phenolic components, and antioxidant activity [
103]. Other important bacteria are
Enterobacter spp.,
Pseudomonas spp.,
Ochrobactrum spp.,
Bacilus spp., and
Rhodococcus spp. [
104,
105,
106,
107,
108,
109,
110,
111,
112,
113].
Figure 1 shows different classifications of plant biostimulants.