1. Introduction
Osteopontin (OPN) is a secreted glycoprotein which was first discovered as a gene involved with activated T-cell responses in the resistance to human scrub typhus, and named an early T-cell activation gene (Eta-1) [
1]. It was also identified in separate studies in bone [
2,
3]. The name, osteopontin, was coined based upon the rat bone sialoprotein to reflect that it is formed in the bone environment (
osteo) and can form a bridge (
pon) between cells and the hydroxyapatite matrix [
4]. Subsequently, intracellular forms were identified and it is expressed in a broad range of tissue and cell types such as hematopoietic, fibroblastic, and immune cells. The expressed levels of OPN can be increased in different cell types by sundry inducers such as by the stimulation of the immune system or by inflammation [
5]. OPN is present in mammalian milk at high levels and is among the top ten most abundant proteins in milk [
6].
In control animals and people without disease, no physiological function has been proven for OPN. Mice with a full deficiency of OPN (OPN-KO) are fertile and undergo normal embryogenesis and development, and do not possess any phenotype before the challenge [
7]. Even in the first report on OPN-KO mice, however, wound healing in response to a skin injury was shown to be modified in the absence of OPN. Detailed studies of pups born to OPN-KO females compared to those born to wild-type (WT) females show that the presence of OPN in milk promotes the development of the brain, intestine, and immunity, and leads to improvements in cognition, immune function, and intestinal development in early life [
8]. When OPN is lacking in milk in OPN-KO mouse pups nursed by OPN-KO mothers, there is a reduced expression of OPN ligands, CD44, and integrins αv and β3 in the jejenum, resulting in lower signaling in the Erk, PI3K/Akt, Wnt, and FAK pathways [
9]. Taken together, these data have resulted in recombinant OPN being evaluated as an additive for human infant nutrition [
10].
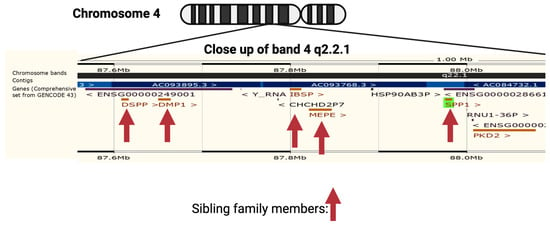
OPN is encoded by a single copy gene on human chromosome 4 (4q13), called the secreted phosphoprotein 1 (
SPP1) gene (
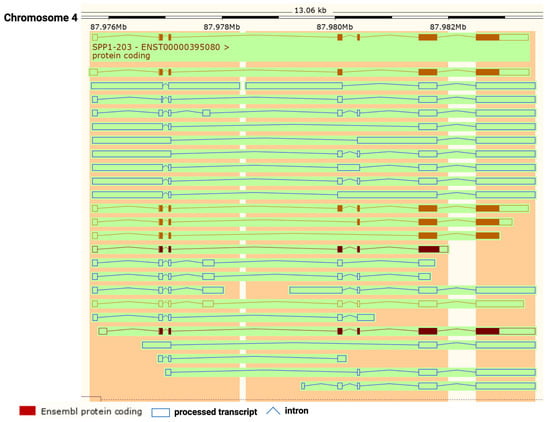
Figure 1). The SPP1 gene contains seven exons [
11] which result in 26 splice variants found in the ENSEMBL database (
Figure 2). Of those 26 OPN mRNAs, only 6 are protein coding and 2 of those give rise to identical full-length proteins (osteopontin-a) that, in humans, consist of 314 amino acids (OPN residue numbering used in this review is based on the full-length human osteopontin-a protein including the signal sequence, unless otherwise specified). The predicted molecular weight of the unmodified peptide chain is 36 kDa but the protein exists in a variety of forms ranging from 41 to 75 kDa depending on the degree of post-translational modification (see below).
Figure 1. Neighborhood of osteopontin (
SPP1) gene on human chromosome 4. The genes encoding members of the small integrin-binding ligands with N-linked glycosylation (SIBLING) family are marked with a vertical arrow: bone sialoprotein (
BSP), dentin matrix protein 1 (
DMP1), dentin sialophosphoprotein (
DSPP), matrix extracellular phosphoprotein (
MEPE), and Osteopontin (
Spp1). Created with
BioRender.com with data from ENSEMBL v109, human assembly GRCh38.p13 [
12].
Figure 2. Osteopontin (OPN) splice variants. Transcripts from
SPP1 gene with protein-coding exons shown in filled-in brown boxes. Other processed transcripts that are not translated are shown as blue open boxes. Introns are shown as blue lines. Created with
BioRender.com with data from ENSEMBL v109, human assembly GRCh38.p13 [
12].
Several transcription factor binding sites have been identified in the OPN
SPP1 promoter including Ets1, β-catenin, runx1, C/EBPα, and AML1, all of which have previously been shown to have tumorigenic roles [
13,
14,
15]. Transcription of the
SPP1 (OPN) gene is also regulated by miR-181a both in glioblastoma as well as during aging [
16,
17]. In addition to the regulation of the OPN expression at the transcriptional level, OPN mRNA is regulated by the interaction of the 5′ UTR with elongation factor 1A1 (EF1A1) [
18].
A large number of polymorphisms have been identified along the
SPP1 gene and some have been associated with cancer and autoimmune diseases such as rheumatoid arthritis, asthma, and inflammatory bowel disease [
19,
20,
21]. In urolithiasis, the OPN polymorphisms might affect the regulation of renal calcification and stone formation [
22,
23]. There are more than 300 variant sequences of the human
SPP1 gene catalogued in the genome databases. Of those variants, about 10 correspond to short deletion and insertion polymorphisms (indels), and the remainder are single nucleotide polymorphisms or variants (SNVs).
Some of the OPN splice variants can be retained intracellularly [
24] and the intracellular forms, osteopontin-c and osteopontin 5, are translated from an initiation codon in an exon accessed by alternative splicing [
25]. Intracellular OPN interacts with BCL-6, a transcription factor, regulating the differentiation of follicular helper T cells [
26]. The intracellular forms of OPN can accumulate in the nucleus where it might serve as a prognostic marker for breast cancer [
27]. Intracellular OPN stabilizes TRAF-3, a component of the TNFR signaling pathway, by preventing its degradation via ubiquitin, leading to enhanced protection from viral infection by the increased production of IFN-b [
28]. Thus, the intracellular forms of OPN can modulate immune system responses but, here, we focus on the role of thrombin cleavage of the extracellular forms of OPN.
Genome-wide association studies (GWAS) have shown a correlation of particular polymorphisms in the promoter region of OPN (−443 and −616) with the metastasis and prognosis of both gastric cancer and hepatocellular carcinoma [
29,
30,
31]. A weak association of OPN SNVs with nasopharyngeal carcinoma has been found [
32]. These studies, however, were carried out by examining registries and samples from the Chinese population and may not be generalizable. The risk of glioma is also related to polymorphisms at −155/−156 insG and −443 within the OPN promoter and those have been proposed for targeted cancer gene therapy [
33,
34]. When these polymorphisms were tested in reporter gene constructs, they all affected transcription from the OPN promoter [
35]. Other SNVs in the OPN promoter as well as elsewhere in the gene have associations with cancer prevalence and some of those SNVs also occur within predicted transcription binding sites [
36,
37].
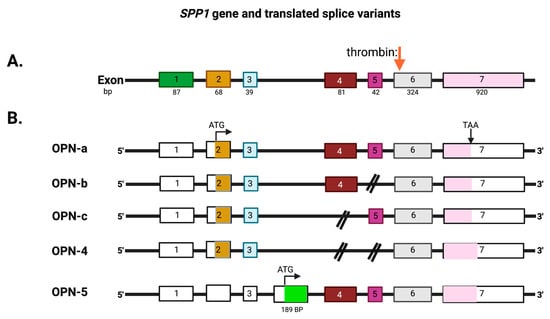
The mammalian
SPP1 gene encodes seven exons that are transcribed into OPN mRNAs of which exon 1 is not translated, while exons 2–7 encode the protein-coding sequences (
Figure 3). Exon 2 encodes the signal peptide (SP) plus the first two amino acids of the mature protein. Exon 3 encodes sequences that can be phosphorylated at the serine residues and exon 4 encodes a proline-rich region and includes two glutamine residues (Q50 and Q52) that are potential transglutamination sites [
38,
39]. Exon 5 encodes another protein phosphorylation site. Finally, exons 6 and 7 encode the remainder of the OPN protein that includes the integrin-binding sites and thrombin and some other protease cleavage sites. A splice variant, osteopontin-b, has been identified that lacks exon 5. The different splice variants including osteopontin-a, -b, and -c, are expressed differentially in cancers [
40,
41]. A meta-analysis based on data in published literature supplemented with data from the TSVdb concluded that in lung cancer, OPN-a, OPN-b, and OPN-c were increased in comparison to normal tissue, while in breast cancer, OPN-c was elevated [
42].
Figure 3. Exons in the osteopontin gene. (
A) OPN (
SPP1) exons on chromosome 4. (
B) The exons that are translated from the OPN mRNAs into the extracellular forms, OPN-a, OPN-b, OPN-c, and OPN-4, and the intracellular form, OPN-5, with its unique exon (in green). The full-length protein of 314 amino acids, OPN-a, is translated from the upper splice variant. Thrombin cleavage site: red arrow. ATG: start codon. TAA: stop codon. Created with
BioRender.com.
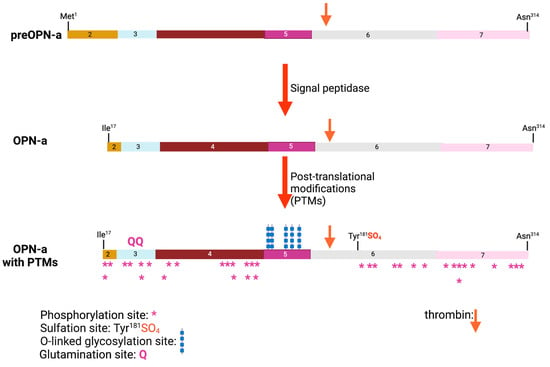
Within the region encoded by exons 6 and 7 is a conserved Arg-Gly-Asp (RGD) sequence that interacts with αvb1, αvb3, and αvb5 integrins, allowing binding to various components of the extracellular matrix as well as some cell types. The adjacent Ser-Val-Val-Tyr-Gly-Leu-Arg (SVVYGLR) sequence domain interacts with α4b1, α9b1, and α4b7 integrins only when this sequence is exposed at the C-terminus of the N-terminal OPN fragment in response to thrombin cleavage. It is functionally inactive in intact full-length OPN (OPN-FL). Thrombin cleavage generates an N-terminal fragment that includes both of these integrin-binding sites and gives rise to a C-terminal fragment containing calcium- and heparin-binding sites, which interacts with some variants of the cluster of differentiation 44 (CD44), such as CD44v3, CD44v6, and CD44v7 [
7,
8] (
Figure 3A and
Figure 4). In addition to thrombin cleavage, OPN can also be cleaved by other proteases including matrix metalloproteinases (MMP)-3 and MMP-7, plasmin, and cathepsin D. The proteolytic cleavages can render these domains more readily accessible, thus enhancing OPN binding efficiency to the integrins. The OPN variants generated by these enzymatic cleavages may retain their activities or acquire or lose additional functions [
9,
10].
Figure 4. Osteopontin post-translational modifications (PTMs). PTMs are marked for glutamination, O-linked glycosylation, phosphorylation, and sulfation, and the thrombin cleavage site is marked with a red arrow. The numbers represent the exon encoding that sequence. Created with
BioRender.com.
Human OPN has conserved a similar domain structure and other features such as the integrin-binding sites and the thrombin cleavage site as other mammalian OPNs. Mammalian OPNs have strong homology over most of the protein sequence with OPNs from reptiles and birds except for sequences close to the C-terminus which, while highly conserved within mammals, are completely distinct in reptiles and birds, suggesting that these C-terminal sequences have unknown but different functions in mammals compared to reptiles and birds [
43]. In addition to these tetrapods, the OPN gene exists in fish, supporting the concept that it was involved in the early evolution of bony skeletons and teeth.
2. The Role of OPN in Cancer
There are several strong lines of evidence that OPN is involved with both primary tumor formation as well as metastasis. Those include the correlation between levels of OPN protein either in blood or in tumor biopsies with worse outcomes; data from single-cell RNA analysis of tumor biopsy samples detail the production of OPN by tumor cells as well as tumor-associated leukocytes, experiments exploring mechanisms upstream from the transcription of the OPN gene (
SPP1) and downstream processes resulting from increased levels of OPN, as well as animal studies investigating the role of OPN in various models of cancer and, in particular, in genetically modified mice. OPN production by the primary tumor and its circulation may cause potential sites for metastasis to be preconditioned especially in the bone [
93,
94]. Taken together, the overall evidence is consistent with a key role for OPN in tumorigenesis and metastasis. A proof that OPN plays an essential function in these processes, however, is still lacking for the human disease. Most of these studies have not distinguished between different forms of OPN and, in general, have not investigated the role of thrombin or other protease cleavages of OPN in cancer (see below).
3. Increased Expression of OPN and Cancer
High serum or plasma levels of OPN have been associated with many human diseases, especially those that involve inflammation. These diseases include cancer in which high levels of OPN are prognostic for worse outcomes in several tumors, and its levels have been suggested as a marker for response to therapy [
95]. Metastasis in melanomas, glioblastomas, and ovarian, lung, and breast carcinomas correlates with increased OPN levels [
96,
97,
98]. The clinical utility of OPN as a marker in cancer therapy has been recognized by its inclusion in test panels for screening for eight tumors via liquid biopsy [
99]. Indeed, when the expression of OPN was investigated in tumor tissues, the OPN gene
SPP1 was in the top 5% of overexpressed genes by microarray [
100]. In several cancers, such as cervical squamous cell carcinoma and endocervical adenocarcinoma, colon adenocarcinoma, brain lower-grade glioma, liver hepatocellular carcinoma, lung adenocarcinoma, and rectum adenocarcinoma, the levels of OPN mRNA levels in tumor biopsies from patients predicted survival (
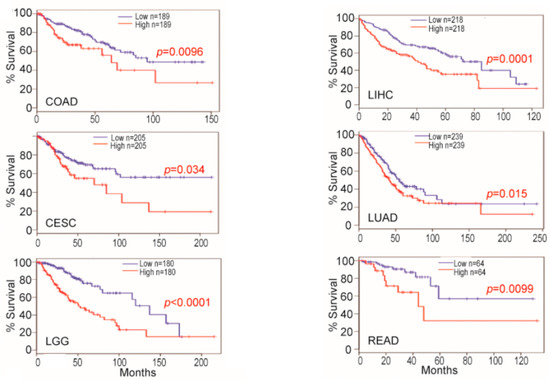
Figure 5) [
101,
102].
Figure 5. Osteopontin (OPN) mRNA levels in non-neoplastic colon and colon carcinoma predict survival. OPN mRNA expression level and survival datasets were extracted and plotted for survival. For each indication, the values considered high and low levels of OPN are shown in ng/mL. Blue line: survival of low OPN group, red line: survival of high OPN group. CESC: cervical squamous cell carcinoma and endocervical adenocarcinoma; COAD: colon adenocarcinoma; LGG: brain lower-grade glioma; LIHC: liver hepatocellular carcinoma; LUAD: lung adenocarcinoma; READ: rectum adenocarcinoma. Data from [
102]. Created with
BioRender.com.
In lung cancer, high blood OPN levels are a prognostic marker for worse outcomes such that patients with high OPN levels had a median survival of 57 months, while those with low OPN levels had a median survival of 102 months (hazard ratio = 1.57 (1.36–1.81),
p = 5.390 × 10
−10) [
97]. OPN is overexpressed in lung cancer tissues and the tissue OPN is also associated with a poor prognosis and decreased survival rates in lung cancer patients. Serum OPN levels can be used as a prognostic marker in small-cell lung cancer and, in combination with CEA levels, as a diagnostic marker in non-small-cell lung cancer [
103,
104]. Overexpression of OPN is associated with poor outcomes in Alk1 fusion lung cancer patients who do not receive targeted therapy [
105]. Plasma OPN levels serve as a better diagnostic marker than mesothelin levels for the diagnosis of mesothelioma [
106]. In patients with lung cancer, circulating levels of OPN correlate with those of VEGF and MMP-9, suggesting that high OPN correlates with increased angiogenesis and, hence, the increased aggressiveness of the tumor [
107].
After surgery in hepatocellular carcinoma patients, high plasma OPN levels correlated with a lower overall survival when compared to patients with low OPN levels (
p = 0.001) [
108]. In a meta-analysis of hepatocellular carcinoma that included 12 studies with 2117 patients published up to 2017, high blood OPN levels were associated with poor overall survival (OS; OR = 1.84; 95% confidence interval [CI] = 1.54–2.20;
p = 0.0001) and disease-free survival (DFS; odds ratio (OR) = 1.67; 95% CI = 1.40–1.98;
p = 0.0001) [
109]. Furthermore, in the subgroup analysis, high levels of OPN in tissues assessed by immunohistochemistry detection and serum-based OPN by ELISA detection were both associated with worse OS (tissue: OR = 1.88; 95% CI = 1.53–2.31;
p < 0.0001; serum: OR 2.38; 95% CI 1.58–3.59;
p < 0.0001). In the same study, OPN expression was positively associated with the stage (OR = 5.68; 95% CI 3.443–7.758) and tumor size (size ≤ 5 cm vs. >5 cm; OR = 2.001; 95% CI = 1.036–3.867). Subsequent studies have confirmed that HCC patients could be stratified based on their OPN levels [
110,
111]. Extracellular vesicles released from a tumorigenic HCC cell line, HepG2, contained more OPN and less miR-181a, an miRNA that down-regulates OPN transcription, than extracellular vesicles from a non-tumorigenic line, WRL68 [
112]. The current evidence indicates that OPN could serve as a prognostic biomarker and a potential therapeutic target for HCC.
High levels of OPN expression are associated with advanced stages of breast cancer, with lymph node metastasis, and with poor prognosis. Levels of OPN expression have been proposed as a predictive biomarker in the anti-EGFR therapy of triple negative breast cancer, but this proposal was based on in vitro experiments on breast cancer cell lines [
113]. Higher OPN protein overexpression is found in breast tumors than in normal breast tissue, and higher plasma levels of OPN are positively associated with increased tumor burden and the shorter survival of patients [
114,
115]. In a meta-analysis of 10 breast cancer clinical studies, including a total of 1567 participants, both high levels of serum and tissue OPN indicated a poor breast cancer outcome. Moreover, OPN expression levels correlate with the expression of several other biomarkers, such as p53, other inflammatory markers, HER2, estrogen receptor (ER), and progesterone receptor (PR) [
116,
117]. In a study on tumor recurrence in tamoxifen-treated breast cancer patients, biopsies were evaluated for OPN mRNA expression by qPCR and OPN protein by immunohistochemistry [
118]. OPN mRNA expression increased the risk of recurrence with an OR = 2.50 (95% CI; 1.30–4.82) for which, when adjusted for tumor grade, the HER 2 status and other treatments had an OR = 3.62 (95% CI; 1.45–9.07). OPN protein expression, however, was not associated with the risk of recurrence or with OPN mRNA expression, suggesting that OPN mRNA is a stronger prognostic marker than tumor tissue OPN protein.
A meta-analysis of ovarian carcinoma patients showed that they had higher levels of serum OPN than in healthy controls [
119]. In this study, a subgroup analysis by ethnicity suggested that high levels of serum OPN might be the main risk factor for ovarian neoplasms in Asians (
p < 0.001), but not in Caucasians (
p > 0.05). Increased OPN mRNA expression levels are also associated with poor prognosis and decreased survival in ovarian carcinoma patients [
120]. In an Indian population from South Asia, when OPN was compared with carbohydrate antigen 125 (CA125) as a diagnostic marker for ovarian carcinoma, OPN had a higher specificity than CA125 in detecting ovarian cancer. OPN can also differentiate better between benign and malignant ovarian cancer than CA125, suggesting its use as a diagnostic marker [
121].
OPN expression levels are significantly higher in cutaneous melanoma patients than in healthy individuals, and higher OPN levels are associated with poor prognosis in melanoma patients [
122,
123]. In uveal melanoma, a proteomics study suggested OPN levels as a potential biomarker [
124]. The V600E BRAF mutation is common in melanomas and is the target for the specific chemotherapy of melanoma. Levels of OPN and MMP9 were correlated and higher in melanoma patients than in controls but, in those patients with the V600E BRAF mutation, the levels of both OPN and MMP9 were not different than in those patients without that mutation [
125]. Resistance to a BRAF inhibitor, the vemurafenib analogue PLX4720, was induced in four paired primary/metastatic cell lines, and, in each case, OPN expression was reduced in a manner similar to that seen in non-small-cell lung cancer [
126,
127]. Follow-up studies on melanoma cells resistant to both a BRAF inhibitor (encorafenib) and an MEK inhibitor (binimetinib) found that OPN was one of several genes whose expression was unaltered during a drug “holiday” [
128]. These data support a role for OPN in the pathogenesis of melanoma and its potential as a marker, possibly to track resistance to chemotherapy.
Glioblastoma multiforme (GBM) is a highly aggressive and lethal brain tumor with poor prognosis, and its treatment remains a challenge for oncologists. OPN is highly expressed in GBM and has been implicated in the pathogenesis and progression of this malignancy such that increased serum levels of OPN correlate with poor prognosis [
129,
130]. OPN is also secreted into the CSF where its thrombin cleavage fragments have been detected [
131,
132]. OPN was one of five genes that distinguish highly invasive incurable glioblastoma from the less aggressive lower-grade astrocytoma in a study using differential expression to identify highly up-regulated genes [
133].
These studies are representative of many others that, together, demonstrate a strong correlation between OPN levels in the blood with poor outcomes and support the use of OPN levels as a diagnostic marker. It also offers the possibility of following therapeutic outcomes in several cancers, but further studies are needed to validate that concept.
In all of these studies correlating OPN levels with outcomes in cancer and other diseases, the assays used to measure OPN levels in blood or tumor biopsies were commercial ELISAs that neither distinguish between the different splice forms of intact OPN-FL nor between the different proteolytically cleaved fragments of OPN. Indeed, in many cases, the exact OPN forms being detected are unknown. Future studies would be enhanced if they employed ELISAs that had been fully characterized with respect to the different forms of OPN being detected.
The role of the thrombin cleavage of OPN has not been investigated extensively in human cancer and, to our knowledge, the only studies in which intact OPN-FL has been specifically distinguished from the thrombin cleavage fragments have been that by Yamaguchi et al. investigating GBM [
132]. Thrombin-cleaved OPN (OPN-arginine or OPN-R) and thrombin followed by basic carboxypeptidase double-cleaved OPN (OPN-leucine or OPN-L) were detected at higher levels in cerebrospinal fluid from cancer patients compared to non-cancer patients, and, in tissue biopsies, glioblastoma had increased OPN-R and OPN-L compared to tissues from patients with epilepsy.
4. OPN mRNA Expression in Tumor Cells and Tumor-Associated Cells
Evidence that OPN mRNA could be up-regulated in cells in tumors was first described in a rat osteosarcoma cell line [
134] and has been confirmed in other cell culture models as well as in animal and human tumor biopsies [
135,
136,
137,
138]. One report demonstrated the role of the Wnt/β-catenin pathway in increasing OPN expression in colon cancer and its consequences on reduced survival when OPN is high [
101]. Other signaling pathways, including hedgehog/Gli, can stimulate OPN mRNA expression [
139,
140].
Bulk RNA sequencing has been used to compare tumors grown in mice with genetic changes, such as a study on the role of NFAT2c in prostate cancer pinpointing the increase in OPN expression [
141]. These studies and subsequent follow-up reports showed that both tumor cells and tumor-associated cells that form the tumor microenvironment could up-regulate OPN mRNA expression, but the precise cell types involved in human cancer were not elucidated.
Single-cell RNA sequence analysis overcomes that limitation and has shown that tumor-associated macrophages (TAMs), in particular, can increase OPN expression, leading to changes in their phenotype as well as influencing the overall outcome of the disease, such as in colorectal carcinoma and pancreatic ductal adenocarcinoma [
142,
143]. Importantly, the data from studies in colorectal cancer and cervical cancer suggest that OPN-producing TAMs may interact with tumor-associated fibroblasts to form the tumor microenvironment [
144]. Use of the single-cell RNAseq technique allowed the demonstration of increasing heterogeneity and worse prognosis in more advanced hepatocellular carcinoma and showed that OPN expression correlated with tumor cell evolution and the reprogramming of the tumor microenvironment [
145]. In gall bladder carcinoma, TAMs expressing OPN were associated with poor prognosis in PD-1-treated patients [
146].
Characterization of TAMs in GBM by an RNA microarray found that they could not be classified into the simple M1 vs. M2 nomenclature [
147]. Instead, the TAMs lay on a continuum between the M0 and M2 types of macrophages. That classification was originally derived from a comparison of macrophage responses that differ between mouse strains [
148]. In this scheme, classic M1 macrophages are defined by the expression of the transcription factor, STAT1, and lead to an anti-tumor response via the presentation of tumor antigens to adaptive immune cells [
149]. In contrast, alternatively activated M2 macrophages express intracellular STAT3 and the scavenger receptors, CD163 and CD204, as well as the mannose receptor, CD206, leading to the secretion of cytokines that suppress the host anti-tumor immune response [
150].
Datasets of single-cell RNAseq from patients with non-small-cell lung cancer (NSCLC) who were smokers were first analyzed to identify clusters of cells using the expression of known markers for cell types, such as tumor cells with EPCAM and leukocytes with PTRPC (CD45) [
151]. Then, the leukocytes were separated into classes using a similar method before splitting the myeloid cells into clusters of different cell types. Two types of immune-suppressive macrophages were found, one characterized by the expression of CCL18 which reduces the production of inflammatory factors, and the other categorized by OPN (
SPP1) expression which promotes angiogenesis and matrix remodeling. Differences between samples from males and females were noted, with samples from males having a higher expression of the matrix proteins, OPN and
FN1, as well as the complement protein,
C1QC. Analysis of these three genes from other cohorts suggested that the higher expression of
FN1,
SPP1, and
C1QC in immune cells correlates with a poorer prognosis. These differences might explain the sex-related difference in the prognosis of NSCLC, that is mainly due to variations in the immune response [
152].
Another study analyzed single-cell RNAseq generated from early-stage NSCLC patients, finding that several cell types comprising a cell module that was named “the lung cancer activation module (LCAM
hi)” could be identified. It consisted of PDCD1
+CXCL13
+ activated T cells, IgG
+ plasma cells, and SPP1
+ macrophages [
153]. These tumor-associated macrophages are derived from monocytes and are distinct from alveolar macrophages. Using immunohistochemistry tumor markers for LCAM
hi cells to evaluate biopsies of NSCLC, this cellular signature was shown to be independent of overall immune cell content but did correlate with the tumor mutational burden, cancer testis antigens, and P53 mutations. High baseline LCAM
hi scores correlated with an enhanced NSCLC response to immunotherapy.
Lymph node metastasis can occur early in lung adenocarcinoma (LUAD) [
154]. Biopsies from 16 patients with LUAD, half with metastasis and the other half without, were studied by single-cell RNAseq [
155]. M0 macrophages were increased in the metastatic group while the epithelial–mesenchymal transition (EMT) pathway was increased in patients with higher M0 infiltration levels. OPN (
SPP1) is also a gene involved in EMT that correlates with macrophage infiltration and M2 polarization. OPN was shown to be a potential marker for early lymph node metastasis in early-stage LUAD.
In glioblastoma, single-cell RNA sequence analysis has shown that tumor-associated fibroblasts are producing OPN [
156]. This signaling network between the tumor cells, TAMs, and tumor-associated fibroblasts is not just present in primary tumors but is also established by colorectal cancer metastases in the liver [
142]. In colorectal carcinoma, combining spatial transcriptomics with a single-cell RNA sequence allowed the colorectal cancer cells to be localized to the invasion front where the expression of HLA-G caused the induction of TAMs. These TAMS then produced OPN, which modified the tumor microenvironment to one with anti-tumor immunity, as well as increasing the potential for the proliferation and invasion of the tumor cells [
157]. This type of network in which OPN acts as part of a communications interaction system between different cell types could be a common feature of cancers.
9. OPN in Tumor Cell Culture Models: Its Expression and Its Effects
Secreted OPN, whether produced by the cell itself and acting in an autocrine fashion or produced by other cells as a paracrine factor, stimulates a variety of effects on cells via the various different cellular receptors for OPN. In the context of tumors, OPN produced by tumor cells will modulate the behavior of those cells as well as cells in the tumor microenvironment. Effects of OPN on the tumor cells include changes in adhesion, migration, apoptosis, and proliferation. Cells in the tumor microenvironment are regulated by OPN to create a more tumorigenic niche. These changes include the generation of tumor-associated fibroblasts from resident fibroblasts and mesenchymal stem cells while inducing an immunosuppressive environment by the modulation of TAMs and regulatory T cells [
158]. OPN also increases angiogenesis, critical for tumor survival and growth.
Studies have shown that OPN promotes lung cancer cell proliferation, invasion, and metastasis through several signaling pathways, including the PI3K/Akt and MAPK/ERK pathways. Overexpression of OPN in prostate cancer PC3 cells as well as non-transformed rat vascular smooth muscle cells led to the phosphorylation of ERK1/2, JNK, and p38 MAPK, three members of the MAPK/ERK pathway [
159,
160]. In human chondrosarcoma cells, OPN increases the phosphorylation of MEK, a MAPK, and FAK, and leads to the overexpression of MMP9, promoting more migration [
160].
In a breast cancer cell line (MDA-MB-231), the knock-down of OPN with lentivirus led to reduced levels of integrin α
vb
3 with the subsequent inhibition of migration and invasion, along with a decrease in signaling in the PI3K/Akt pathway and an increase in autophagy-related proteins (LC3 and Beclin 1) [
161]. OPN overexpression in prostate cancer epithelial cells (PC3, DU145, and LNCaP) leads to the activation of Akt via PI3K and ILK, which then regulates the expression of b-catenin [
162].
OPN also contributes to the development of the resistance to both classical chemotherapy and targeted therapies in lung cancer. The inhibition of OPN expression or activity sensitizes lung cancer cells to chemotherapy and enhances the efficacy of targeted therapies such as EGFR inhibitors [
163,
164]. In contrast, one study in lung adenocarcinoma reported that OPN could cause EGFR phosphorylation, leading to sensitization to EGFR inhibitors [
165].
OPN has also been found to interact with other signaling pathways and molecules that are involved in GBM, such as the Akt/mTOR pathway, matrix metalloproteinases (MMPs), and integrins. For example, OPN can activate the Akt/mTOR pathway and induce the epithelial–mesenchymal transition (EMT), a process that promotes cancer cell invasion and metastasis [
166]. OPN can also induce MMP expression and activity, which, in turn, can degrade the extracellular matrix and promote tumor cell invasion and angiogenesis. When OPN is bound to integrins, such as α
vβ
3 and α
vβ
5, it can promote malignant astrocytoma cell adhesion and migration [
167].
The addition of exogenous OPN protein to cell cultures changes the cell behavior in many functional assays such as ones for adhesion, apoptosis, migration, and proliferation. These effects are similar to those caused by the expression of OPN. Treatment of human mammary epithelial cells with exogenous OPN caused an increased induction of uPA expression, accompanied by an increase in migration [
168]. In four human colon cancer cell lines, adding OPN caused a reduction in intertypic cell adhesion, an increase in migration via CD44, and an increase in invasion into Matrigel when OPN was included as a chemoattractant [
169]. When these cell lines were transfected with a plasmid-encoding OPN and grown as subcutaneous tumors in nude mice, there was an increase in VEGF production in the tumor and more metastasis.
When HCC cell lines were exposed to different splice variants of OPN, OPN-a, OPN-b, and OPN-c, a migratory line, Hep3B, increased its migration in response to OPN-a and OPN-b but not to OPN-c, while in a non-migratory line, SK-Hep1, OPN-a had no effect and OPN-c inhibited migration [
170]. In HepG2 cells, exogenous NO stimulates transcription from the OPN promoter and NO is also produced in the tumor microenvironment both by hepatoma cells and macrophages [
171,
172,
173].
Angiogenesis is enhanced by the presence of OPN, resulting in the increased production of angiogenic factors such as VEGF or PGE
2 by tumor cells and the stromal cells in the tumor microenvironment, as well as having direct effects on endothelial cells. The role of OPN in inducing VEGF and, subsequently, angiogenesis was demonstrated in both breast and colon cancer cells by siRNA knock-down [
174,
175]. In prostate cancer cells, OPN and MMP9 expression result in enhanced VEGF production [
176]. Hypoxia is a common feature of solid tumors, leading to the induction of a large number of different proteins commonly via HIF1α. In breast cancer, OPN mRNA is induced by hypoxia, but its induction was independent of HIF1α [
175]. The increased OPN leads to a positive feedback loop in which ILK and Akt are activated, causing the production of VEGF.
Endothelial precursor cells respond to the overexpression of OPN by accelerating angiogenesis in vitro via increased proliferation, migration, and tube formation. This is mediated by OPN interacting with integrin α
vb
3 on endothelial precursors, thereby leading to the activation of PI3K, Akt, and eNOS that, downstream, produce higher levels of NO. In mature endothelial cells, OPN treatment activates both the PI3K/Akt and the ERK1/2 pathways, inducing angiogenesis [
177].
14. Therapies Targeting OPN in Cancer
Given the data above, the cleavage of OPN by thrombin is a target for cancer therapy. Several approaches are possible for defining the precise target for the treatment, including inhibiting thrombin cleavage of OPN with anticoagulants, blocking the activities of either OPN-R or OPN-CTF, and antagonizing specific binding domains within OPN.
Anticoagulation has been studied as a treatment for cancer, including trials on warfarin and heparin. There is also a significant risk of thromboembolic events in cancer patients, and treatment as well as secondary prevention and low-molecular-weight heparin was used for these purposes. In addition, some cancer patients had previously been diagnosed with atrial fibrillation and prescribed warfarin for stroke prevention. Because of the lack of specificity of these earlier agents and the known disadvantages of i.v. or s.c. dosing for heparin and low-molecular-weight heparin [
222], respectively, and the problem of maintaining consistent anticoagulant effects with warfarin and its low therapeutic ratio, trials of these agents were inconclusive. Argatroban, a direct thrombin inhibitor with low specificity, was shown to reduce the growth, invasiveness, and metastasis of a human breast cancer cell line in mice [
223]. In preclinical studies, hirudin, a parenteral direct thrombin inhibitor, prevented metastasis of B16 melanoma cells in mice [
224]. The newer direct oral anticoagulants, such as the thrombin inhibitor dabigatran and the factor Xa inhibitors, rivaroxaban and apixaban, circumvent those problems but the data so far in cancer in patients who do not have therapeutic or prophylactic indications for treatment with anticoagulants are not definitive [
225].
The survival benefit of direct oral anti-coagulants (DOACs) over warfarin or heparin in cancer patients is unproven in clinical trials so far, maybe because of the sample sizes in the trials being too low and not allowing sufficient time for follow-up [
226]. Currently, there are ongoing population-based studies suggesting that warfarin may be associated with improved overall survival in cancers and may protect against certain cancers. The anti-tumor effects of warfarin could occur through both coagulation-pathway-dependent and -independent mechanisms mediated by the inhibition of the Gas6-AXL signaling pathway. However, the use of anticoagulants to treat cancer patients is limited by the risks of bleeding. It is well-established that cancer patients have a higher incidence of major bleeds (6.5–18%) than patients without cancer (2–3%) on anticoagulation [
227]. This increased bleeding cannot be fully accounted for by supratherapeutic anticoagulation and, instead, is likely to be related to abnormal tumor vasculature characterized by its immaturity and “leakiness” [
228,
229]. In addition, anticoagulation may be contraindicated in patients with primary brain cancers, such as GBM, and cancer patients with intracerebral brain metastasis, because of the risk of catastrophic intracranial bleed [
230,
231].
Anti-OPN mABs have been identified that bind different parts of the OPN polypeptide sequence and have been tested in preclinical cancer models and in the clinic for other indications. An mAB directed against the novel C-terminal site of OPN-R that is formed after thrombin cleavage, administered to a mouse model of human adult T-cell leukemia, resulted in the inhibition of tumor growth as well as tumor invasion and metastasis [
232]. This antibody had a larger effect on tumor growth than an anti-OPN mAB that binds to the RGD sequence, while using both antibodies together had an additive effect.
The humanization of an mAB that binds to the sequence
212NAPSD
216 adjacent to the calcium-binding domain of OPN was characterized in vitro to block the cell adhesion, migration, invasion, and colony formation of MDA-MB-435S cells, a human breast cancer cell line [
233]. When tested in a murine breast cancer xenograft model, this mAB reduced the growth of the primary tumor and inhibited spontaneous metastasis to the lung, liver, and brain.
An anti-OPN mAB was identified that binds to the
162SVVYGLRSKS
171 sequence which is next to the RGD sequence and spans the thrombin cleavage site of OPN [
234]. This mAB blocked OPN binding to integrin α
vb
3, inhibited thrombin cleavage of OPN-FL, and reduced cell migration. Its efficacy was demonstrated in vivo using a model of NSCLC in which the mAB, whether as a single agent or in combination with carboplatin, significantly inhibited the growth of large metastatic tumors in the lung.
Four anti-OPN mABs were identified based on their ability to block the OPN inhibition of T-cell activation [
102]. Two of those clones increased the efficacy of tumor-specific CTLs in killing colon tumor cells in vitro, while, in vivo, they could suppress the metastasis of CT26 colon tumor cells in mice.
Anti-OPN antibodies have been tested in the clinic but, so far, with little success. ASK8007 is a humanized mAB that was derived from a murine mAB, 2K1, and is directed against the new C-terminus of OPN-R, SVVYGLR [
235]. ASK8007 is similar to 2K1 in inhibiting OPN binding to the integrins αvb1, αvb3, αvβ5, α4β1, and α9β1, that interact with RGD and SVVYGLR sequences, respectively. A version of 2K1, C2KI, that is a chimera between the mouse 2K1 Fab domains and human Fc domain, cross-reacts with monkey OPN and treats collagen-induced arthritis in a cynomolgus monkey with no notable toxicological findings [
236]. In a double-blind randomized phase IIA trial, rheumatoid arthritis patients received either ASK8007 or a placebo [
237]. It was safe and well-tolerated but there was no clinical response and no changes in synovial sublining macrophages, although ASK8007 could be detected in the synovial fluid.
Use of mABs targeting OPN may have pharmacokinetic problems, resulting in the need for unrealistically high doses of the therapeutic antibody being required to reach efficacy because of the speed of the turnover of OPN in blood [
238]. This study used heavy atom metabolic labeling to determine the half-life of circulating OPN in the blood of three volunteers and showed that the predicted t
1212 is 11 min. The pharmacokinetic/pharmacodynamic relationship was used to calculate the required dosing of anti-OPN mABs based on assumptions that the mean blood levels of OPN would be 444 ng/mL with a range of 112–1740 ng/mL [
239] and the mAB would be required to achieve a 90% reduction in circulating OPN. The study used four different assumptions about the mAB properties, one with standard mAB properties and the other three that had been engineered in different ways to improve the mAB pharmacokinetics (PK) and pharmacodynamics (PD). Although the calculated dosing regimens are theoretically possible with the most favorable assumptions, the authors’ conclusion was that the high amount of drug substance and inconvenience of administration would generally preclude the type of dosing regimen necessary.
This entry is adapted from the peer-reviewed paper 10.3390/cancers15133480