Cancer research has prioritized the study of the tumor microenvironment (TME) as a crucial area of investigation. Understanding the communication between tumor cells and the various cell types within the TME has become a focal point. Bidirectional communication processes between these cells support cellular transformation, as well as the survival, invasion, and metastatic dissemination of tumor cells. Extracellular vesicles are lipid bilayer structures secreted by cells that emerge as important mediators of this cell-to-cell communication. EVs transfer their molecular cargo, including proteins and nucleic acids, and particularly microRNAs, which play critical roles in intercellular communication. Adipocytes, a significant component of the breast stroma, exhibit high EV secretory activity, which can then modulate metabolic processes, promoting the growth, proliferation, and migration of tumor cells.
1. Introduction
Recent advances in cancer biology have highlighted the significance of understanding the crosstalk between tumor cells and their neighboring microenvironment to unravel the mechanisms governing tumor growth and metastasis [
1]. The tumor microenvironment (TME) comprises all the non-cancerous host cells in the tumor, including fibroblasts, endothelial cells, adipocytes, and adaptive and innate immune cells, as well as its non-cellular components, which include the extracellular matrix (ECM) and soluble products such as chemokines, cytokines, growth factors, and extracellular vesicles (EVs) [
2].
During tumor growth, tumor cells dynamically interact with TME components, contributing to cancer cell survival, local invasion, and metastatic dissemination. For example, endothelial cells promote angiogenesis to counteract the hypoxic and acidic TME, ensuring an adequate oxygen and nutrient supply while removing metabolic waste [
2]. Additionally, cancer-associated adipocytes exchange cytokines and lipids with tumor cells, leading to metabolic rewiring and the acquisition of proinflammatory and invasive phenotypes [
3]. These interactions (among others), play essential roles in orchestrating the hallmarks of cancer [
4], and underscore the importance of intercellular communication within the TME in their maintenance and promotion, which in the long term can promote tumor progression and metastasis [
1].
Extracellular vesicles (EVs) have emerged as key mediators of intercellular communication. EVs are lipid bilayer-enclosed membranous structures secreted by all cells. They can be broadly classified into three subtypes: exosomes, microvesicles, and apoptotic bodies, depending on their biogenesis [
5], and large (lEVs) and small (sEVs) EVs based on their size [
6]. EVs carry diverse biomolecules, including proteins, lipids, and nucleic acids [
7], which are necessary for cell-to-cell communication via cell–surface interactions [
8]. This intercellular trafficking can occur in the form of paracrine signaling and/or with distant cells as a form of endocrine signaling [
9]. Therefore, EVs can elicit pleiotropic responses in recipient cells during both physiological and pathological processes [
10].
Of particular interest among the cargo components of EVs are microRNAs (miRNAs), a class of small non-coding RNAs that function in post-transcriptional regulation of gene expression. They are powerful regulators of various cellular activities including cell growth, differentiation, development, and apoptosis [
11]. This highlights the importance of studying their role in cell-to-cell communication, particularly in pro-tumorigenic processes associated with the TME. Dysregulation of the cellular miRNA profile favoring the expression of tumor-promoter miRNAs (oncomiRs) and downregulating the expression of tumor-suppressor miRNAs have been widely associated with the acquisition of tumorigenic and pro-metastatic capacities [
12,
13]. Several studies have associated cellular miRNA dysregulation with enhanced tumor angiogenesis and vascular permeability [
14,
15], epithelial-mesenchymal transition (EMT) [
16,
17,
18,
19], and recently, adipose tissue remodeling [
20,
21,
22]; processes that favor tumor growth, progression, and metastasis. However, the transport and delivery of miRNAs contained in EVs (EV-miRs) secreted by cells in the TME and their specific roles in the promotion of tumorigenic and pro-metastatic capacities have not been completely elucidated, and it is certainly an exciting and important research field.
2. Adipocytes as an Important Source of EVs in the TME
In cancer development, the tumor-cell intrinsic factors play an important role, however, interactions between the TME and tumor influence the biology and progression of the disease to a large extent as well [
85]. Adipose tissue is composed of adipocytes, stromal vascular fraction, and pluripotent stem cells [
86]. Adipocytes from different adipose tissues have metabolic and endocrine functions such as energy storage, release of free fatty acids, and secretion of adipokines, which are bioactive molecules including cytokines, hormones, and enzymes [
87]. In addition to the classical adipokines, adipocytes produce and release EVs, which have emerged as crucial mediators of adipocyte intercellular communication [
88].
Connolly et al., characterized EVs secreted pre- and post-adipogenesis in a 3T3-L1 cells model and found that production of EVs per cell are greater prior to adipogenic differentiation [
89]. Also, they evidence differences in the composition of EVs secreted pre- and post-adipogenesis, but the functional effects of these EVs are poorly understood [
89]. In a further investigation, the EVs secreted by adipocytes were quantitatively and qualitatively characterized through morphological and size parameters, sucrose flotation properties, protein patterns, and lipidomic analysis [
90]. 3T3-L1 cells were differentiated for EVs production, and isolation by ultracentrifugation resulted in two distinct subpopulations: large extracellular vesicles (lEVs) and small extracellular vesicles (sEVs). Specific protein signatures were identified in these subpopulations of EVs, and lipidomic analysis revealed specific cholesterol enrichment in sEVs. In addition, they confirmed these results in primary murine adipocytes [
90].
Following the characterization of EVs secreted by adipocytes (ADEVs), interest in the functional study of these vesicles has increased. Heretofore, research in ADEVs has demonstrated that they may play an endocrine and paracrine role in health and disease, due to the content of the EVs [
91]. The packaging of proteins and miRNAs into ADEVs can be selective and reflect the microenvironment of the cell; for example, ADEVs released under disease conditions can exacerbate or drive pathologies associated with disease complications [
91]. Recently, adipose tissue has been found as an important source of miRNAs in EVs [
92]. This is evidenced by a substantial decrease in the levels of circulating exosomal miRNAs in an adipose-tissue-specific knockout of miRNA-processing enzyme Dicer mouse model, as well as in humans with lipodystrophy [
92].
Microarray analysis of ADEVs secreted by differentiated 3T3-L1 cells revealed 143 miRNAs, among which miRNAs involved in adipose differentiation, such as let-7b, miR-103, miR-146b, and miR-148a, were significantly upregulated in differentiated cells [
93]. One study on mesenchymal stromal/stem cells (MSCs) from porcine adipose tissue characterized the miRNA expression profile in EVs using RNA-seq and found four annotated miRNAs (miR148a, miR532-5p, miR378, and let-7f) predicted to modulate genes involved in apoptosis, angiogenesis, proteolysis, stem cell differentiation, cellular reprogramming, and cell growth [
94].
A study of exosomes isolated from visceral and subcutaneous adipose samples collected from obese and lean patients revealed 88 mature miRNAs that were differentially expressed and 55 annotated miRNAs in obese visceral samples compared with lean visceral samples. They focus on miRNAs validated or predicted to target miRNAs in TGF-β and Wnt/β-catenin pathways (miR-23b, miR-148b, miR.182, miR-3681, miR-4269, miR-4319, and miR-4429) and finally they evaluated the uptake of exomes in lung epithelial cells. Nevertheless, to fully understand the function of miRNAs in ADEVs more studies will be needed [
95].
The role of adipose tissue in tumor progression has been described and miRNAs contained in ADEVs can modulate fundamental biological capabilities for the development and progression of cancer, such as migration, metabolic reprogramming, angiogenesis, invasion, apoptosis, immune function, and proliferation [
96]. For example, in the regulation of hepatocellular carcinoma (HCC), ADEVs can transfer miR-23a/b to hepatocellular cancer cells and promote cellular growth and migration [
97]. The most recent study by Mathiesen et al., determine the contribution of omental (OM) and subcutaneous (SC) adipose tissue EVs (ATEVs) in metastatic prostate cancer (PC3ML cells) and evaluated the miRNA cargo with NanoString array. Forty-eight miRNAs were detected in SC ATEVs, 55 in OM ATEVs and 44 in both; transcriptomic analyses on PC3ML after treatment with ATEVs showed differentially expressed genes associated with proliferation, invasion and glycolysis and were validated with functional assays [
98].
Studies on the interactions between adipose tissue and tumor cells have gained interest because some tumors start or metastasize near adipose tissue, and breast, prostate, and ovarian tumors are in sustained contact with adipose tissue [
99]. Adipocytes surrounding cancer cells undergo phenotypic changes and are referred to as cancer-associated adipocytes (CAAs) [
90,
100]. CAAs secrete different soluble factors and extracellular vesicles that promote tumor progression [
101,
102,
103]. Currently, there is limited evidence regarding EVs and EV-miRs secreted by CAAs. Au Yeung et al. performed the first study, using RNA-seq to identify higher levels of miR-21 in exosomes from CAAs derived from omental tissues of patients with high-grade serous ovarian cancer than in exosomes isolated from cancer-associated fibroblast tissue. Functional studies also revealed that miR-21 is transferred from exosomes to ovarian cancer cells and confers chemoresistance [
101]. The miRNAs contained in EVs secreted by adipocytes are summarized in
Table 1.
Table 1. miRNAs in extracellular vesicles secreted by adipocytes.
| miRNAs |
Source of EVs |
Description |
References |
miR-148a
let-7f
miR-532-5p
miR-378 |
Porcine adipose-derived stem cells |
Characterized the cargo of EVs by high-throughput RNA sequencing. At least 386 annotated miRNAs were read but four were enriched in EVs. |
[94] |
miR-23b
miR-148b
miR-182
miR-3681
miR-4269
miR-4319
miR-4429 |
Visceral and subcutaneous adipose samples from obese and lean patients |
Compared miRNA levels between obese and lean visceral exosomes. |
[95] |
| miR-31 |
Human ADSCs |
miR-31 promote angiogenesis in HUVECs by targeting factor-inhibiting HIF-1 (FIH1). |
[104] |
| miR-21 |
Normal and cancer-associated adipocytes from ovarian cancer patients |
miR-21 is transferred from cancer-associated adipocytes to cancer cells and confers chemoresistance. |
[101] |
| miR-450a-5p |
Rat adipose tissue and ADSCs |
miR-450a-5p mediates adipogenic differentiation. |
[105] |
| miR-132 |
Human adipose-derived stem cells (ADSCs) |
miR-132 was transferred from ADSCs to lymphatic endothelial cells and promoted proliferation, migration, and tube formation. |
[106] |
| miR-23a/b |
3T3-L1 cells, serum, and tumor tissues of hepatocellular carcinoma patients. |
miR-23a/b was upregulated in serum exosomes and tumor tissue. Results suggested that miR-23a/b was derived from adipocytes and transported into cancer cells, conferring chemoresistance. |
[97] |
| let-7 |
Human ADSCs |
Human ADSC-EVs contribute to angiogenesis via let-7. |
[107] |
let-7i-5p,
let-7f-5p |
Human ADSCs |
Human ADSC-EVs promote migration and invasion of endothelial cells. |
[108] |
| miR-93, miR-125, miR-16, let7, miR-612, miR-494, miR-181 |
Human adipose tissue |
miRNAs contained in ADEVs upregulate genes that may impact increased proliferation and deregulate genes that reduce invasion of prostate cancer cells. |
[98] |
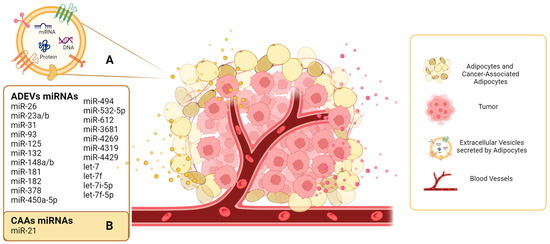
To date, most research has focused on EV-miRs secreted by adipocytes, and there is only one study on EV-miRs secreted by CAAs (Figure 1).
Figure 1. Extracellular vesicles secreted by adipocytes and miRNA cargo. (A) The extracellular vesicles secreted by adipocytes (ADEVs) are represented in yellow, and the miRNAs described to date are shown in the yellow rectangle. (B) In the context of the tumor microenvironment, these adipocytes are transformed into cancer-associated adipocytes (CAAs), and the presence of miR-21 in EVs secreted by CAAs has been described.
The primary cellular component in breast stroma is adipocytes, accounting for 56% of the non-lactating tissue [
109]. In the BC microenvironment, due to the proximity between adipocytes and tumor cells, adipocytes and CAAs are involved in BC progression, and interest in understanding their mechanisms in BC has increased in recent years. To date, many studies have focuses on EVs secreted by adipocytes; however, further studies are required to characterize EVs secreted by CAAs and their miRNAs cargo to understand their potential role in mediating cell-cell communication in the TME in BC.
This entry is adapted from the peer-reviewed paper 10.3390/ijms241713085