1. Tocochromanol Structure
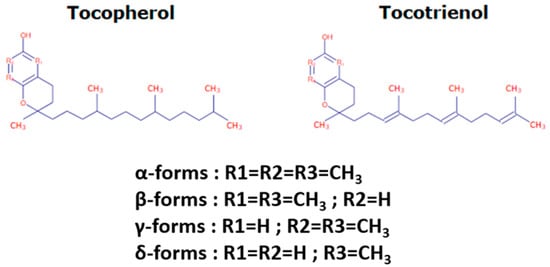
Tocochromanols encompass tocopherols and tocotrienols and are the main components of the vitamin E family. They are exclusively synthesized by photosynthetic eukaryotes and other oxygenic photosynthetic organisms, such as cyanobacteria, and are essential phytonutrients for mammals. In addition, whereas tocopherols are widely distributed in higher plants, tocotrienols occur only in some non-photosynthetic tissues including seeds, roots, and tubers [
24]. Their general structure (reported in
Figure 1) consists of a polar chromanol ring and a hydrophobic 16-carbon side chain attached to the ring via the C-2 atom. While tocopherols have fully saturated 16-carbon phytol side-chains, tocotrienols contain geranylgeranyl side chains with three double bonds in positions C-3, C-7, and C-11. As indicated in
Figure 1, both tocopherols and tocotrienols can occur as four isomers (α, β-, γ-, and δ-) that differ from each other by the number and position of methyl groups in the chromanol ring. The α-isomers are tri-methylated in positions 5, 7, and 8, β- and γ-tocochromanols are di-methylated in position 5 and 8 or in position 7 and 8, respectively, and δ-isomers are methylated in position 8. The chromanol ring forms the basis for the high antioxidant potency of tocochromanols, this potency being modulated by the structure and length of the isoprenoid chains. Actually, tocochromanols are acknowledged as the lipid-soluble antioxidant metabolites possessing the highest capacity to scavenge free radicals [
25]. This is particularly true for α-isomers, which show the highest antioxidant activity among tocochromanols.
Figure 1. Tocochromanol structure.
2. Tocochromanol Biosynthesis
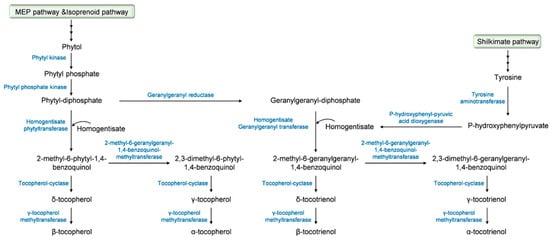
The biosynthetic pathway of tocochromanols has been well documented and we strongly encourage the readers to seek out detailed information in comprehensive studies that have been previously published [
24,
26,
27,
28,
29,
30]. A summary description of this biosynthetic pathway, which starts in the plant cytoplasm for the firsts steps and then takes place in the plastids [
31], is provided in
Figure 2. The precursor of the tocochromanol biosynthesis is the aromatic head group homogentisate that derives from the catabolism of tyrosine into p-hydroxyphenylpyruvate under the action of p-hydroxyphenyl pyruvic acid dioxygenase. The biosynthesis starts with the condensation of homogentisate with different polyprenyl pyrophosphates that determine the type of tocochromanol, phytyl-diphosphate for tocopherols and geranygeranyl-diphosphate for tocotrienols. Phytyl-diphosphate derives from the methylerytrithol phosphate pathway, more precisely from phytol through the successive action of a phytyl kinase and a phytyl phosphate kinase. Another pathway leading to the production of phytyl-diphosphate is the phytol-recycling pathway derived from the chlorophyll degradation [
32,
33]. Geranylgeranyl diphosphate is produced from phytyl-diphosphate
via a reaction catalyzed by the geranylgeranyl reductase. Prenylation of homogentisate with phytyl-diphosphate that leads to the formation of 2-methyl-6-phytyl-1,4-benzoquinol is allowed by the activity of the homogentisate phytyl transferase. For tocotrienols, the prenylation of homogentisate with geranylgeranyl diphosphate is catalyzed by the homogentisate geranylgeranyl transferase that is only located in non-photosynthetic tissues and results in the yield of 2-methyl-6-geranylgeranyl-1,4-benzoquinol [
31]. 2-methyl-6-phytyl-1,4-benzoquinol is acted upon by the tocopherol-cyclase to yield δ-tocopherol or by the 2-methyl-6-phytyl-1,4-benzoquinol-methyl transferase, leading to the formation of 2,3-dimethyl-6-phytyl-1,4-benzoquinol and the tocopherol-cyclase to form γ-tocopherol. The final step of tocopherol biosynthesis involves the methylation of δ- and γ-tocopherols to their β- and α- isomers, respectively, under the effect of the γ-tocopherol methyltransferase [
34]. α-, β-, δ- and γ-Tocotrienols derive from 2-methyl-6-geranylgeranyl-1,4-benzoquinol, which undergoes the same sequence of methylation and cyclization as 2-methyl-6-phytyl-1,4-benzoquinol with the involvement of the 2-methyl-6-geranylgeranyl-1,4-benzoquinol-methyltransferase, the tocopherol-cyclase, and γ-tocopherol methyltransferase.
Figure 2. Tocochromanol biosynthetic pathway.
Knowledge regarding the key genes involved in tocochromanol biosynthesis has initially focused on
Arabidopsis thaliana, being facilitated by the access to
Arabidopsis mutant collections and the possibility to use transgenic plants [
35]. These key biosynthetic genes are reported in
Table 1. In various cereal crops, including maize, barley, and rice, linkage analysis studies of natural variation for tocochromanol levels have led to the identification of QTLs containing tocochromanol biosynthetic genes in their support intervals [
36]. Many of these studies were conducted with a common objective, i.e., improving breeding strategies to increase tocochromanol contents and enriching vegetable oils in vitamin E.
Thus, a genome-wide association approach allowed Schuy et al. [
37] to identify two key genes in barley, the homogentisate phytyltransferase (
HPT) and the homogentisate geranygeranyltransferase (
HGGT) genes; these two genes were shown to be located on chromosome 7H. In maize, candidate genes encoding the core tocochromanol pathway (
ZmVTE genes) have also been characterized [
28]. Regarding oats, the coding sequences of three genes (
HPPD,
VTE2 and
VTE4) have been elucidated [
38]. In rice, biosynthetic genes were first identified by comparative genomics with
Arabidopsis thaliana by Chaudary and Khurana [
39], and the function of a few of them (e.g.,
OsGGR1 and
OsGGR2) was further validated using a transgenic approach [
40]. The former studies were completed by QTL analyses that have led to clarifying the location of
OsγTMT—the gene encoding the γ-tocopherol methyltransferase—on chromosome 2 [
41].
In addition to genes involved in the biosynthetic pathway, genes or genomic areas associated with the natural variations of tocochromanols have been identified. As an example, the gene
LIL3 coding for a light-harvesting-like protein involved in the stabilization of the geranylgeranyl reductase enzyme is a key actor of the genetic control of tocochromanols in
Arabidopsis thaliana [
42]. Significant insights have also been recently published regarding the genetic determinants that govern the accumulation of tocochromanols in maize; more than 40 genomic areas that do not carry the biosynthetic genes have been related to tocochromanol variations [
36]. Among candidate genes identified in the latter study and further supported by the findings of Zhan et al. [
43], homologs of protochlorophyllide reductase (
por) genes (
por1 and
por2) have been shown to account for large allelic effect for tocochromanol traits. These two genes are known to be involved in the regulation of chlorophyll biosynthesis and their association with tocochromanol variations in maize kernels, i.e., kernels that do not contain chlorophyll, first raised numerous questions. These questions were however rapidly resolved with the demonstration that developing maize embryos contain low levels of chlorophyll [
36]. In addition to
por genes, genes coding for plastid-localized fibrillins and cytosolic glycol lipid transfer and SNARE proteins located on four QTLs (QTL10, QTL30, and QTL6) were also suggested to be responsible for tocochromanol phenotypic variation in maize kernels [
36]. In addition, a recent study has indicated the occurrence of genes of the fatty acid pathway in several maize QTLs related to tocochromanol accumulation, suggesting the occurrence of a crosstalk between tocochromanol and fatty acid pathways [
44].
3. Tocochromanol Composition of Major Cereal Crops
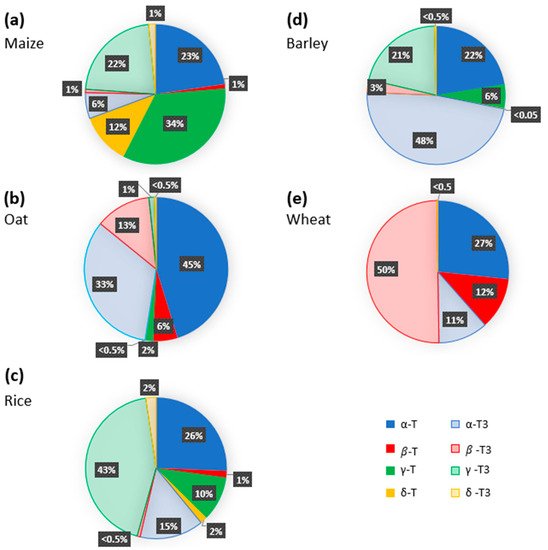
Numerous studies have addressed the tocochromanol composition in kernels of the main cereal types and analyzed the eight tocochromanol isomers, α-, β-, γ-, and δ-tocopherols and tocotrienols. Data from the most recent publications focusing on oat, wheat, barley, rice and maize kernels are gathered in
Figure 3 and
Table S1. Before going further in the discussion, it is important to underline that the mean values reported in
Figure 3 were calculated using different published reports that considered one or a few genotypes and one or a few years of cultivation in some specific agroclimatic conditions. Considering the significant impact of environmental conditions on tocochromanol levels in cereals [
52] and of the genetic background of the cultivar, these data must be taken with caution [
28,
36,
53]. On
Figure 3, it appears that all tocochromanol isomers were found in maize, oat, and rice kernels, whereas γ- and δ-tocopherols and γ-tocotrienol in wheat and β-tocopherol in barley were not detected. The highest levels of tocochromanols, 161 µg.g
−1 dry matter, were reported by Gutierrez-Gonzales et al. in oat kernels [
38]. In wheat, rice, maize, and barley kernels, the highest concentration quantified were 69, 60, 41, and 32 µg.g
−1 dry matter, respectively (
Table S1) [
52,
54,
55,
56]. When considering the relative distribution between tocotrienols and tocopherols, the tocotrienol/tocopherol ratio (calculated with data presented in
Figure 3 and
Table S1) was close to 1 for oat indicating equivalent concentrations of tocotrienols and tocopherols and higher than 1 for rice and wheat, reaching 2.5 for barley. In contrast, maize kernels were shown to contain more than 2 times more tocopherols than tocotrienols.
Figure 3. Tocochromanol profile of major cereal crops ((
a) Maize; (
b) Oat; (
c) Rice; (
d) Barley; (
e) Wheat). The percentages were calculated using average concentration of tocochromanols reported in previously published studies [
38,
52,
54,
56,
57,
58,
59,
61,
62,
63,
65,
66,
67,
68].
The levels of each tocopherol and tocotrienol isomer quantified in whole kernels vary according to the cereal type (
Figure 3). In maize kernels, γ-tocopherol, α-tocopherol, and γ-tocotrienol were the major tocochromanols, which correspond to 34, 23 and 22% of the total tocochromanols (
Figure 3a), respectively, and β-tocotrienol, β-tocopherol, δ-tocotrienol, δ-tocopherol, and α-tocotrienol account for 1 to 12% of the total tocochromanol content [
56,
57,
58,
59]. A different distribution pattern was observed in small-grain cereals. In oat kernels, α-tocopherol and α-tocotrienol were the prevalent tocochromanols, representing 45% and 33% of the total tocochromanol content, respectively, followed by β-tocotrienol (13%), β-tocopherol (6%), and γ-tocopherol (2%) (
Figure 3b) [
60,
61,
62]. The presence of δ-tocotrienol, γ-tocotrienol, and δ-tocopherol was reported as very weak, accounting for less than 1% of the total tocochromanol content. As for oats, the kernels contain the eight tocochromanol isomers, their distribution and concentration being, however, significantly different. In rice, the major tocochromanols are γ-tocotrienol and α-tocopherol, representing, respectively, 43% and 26% of the total tocochromanol content, followed by γ-tocopherol and α-tocotrienol. The sum of δ-tocotrienol, β-tocotrienol, β-tocopherol, and δ-tocopherol accounts for around 5% of the total rice tocochromanol content (
Figure 3c) [
55,
61,
63,
64,
65,
66]. In barley kernels, α-tocotrienol is the main compound and accounts for almost half (48%) of the total tocochromanols [
52,
61,
62]. Both α-tocopherol and γ-tocotrienol were estimated to account to 22 and 21% of the total tocochromanol content, respectively (
Figure 3d). Regarding wheat kernels, the data reported in
Figure 3e indicate that the predominant tocopherol is β-tocotrienol (50%), followed by α-tocopherol (27%) and both β-tocopherol and α-tocotrienol, each of them representing around 11% of the total tocochromanol content [
52,
54,
61,
62,
67,
68].
4. Distribution of Tocochromanols within Cereal Kernels
Enzymes involved in the biosynthesis of tocopherols are located in the inner membrane of chloroplast envelope and in plastoglobuli [
69]. Chloroplasts were first indicated as the unique plant cell organelle containing tocopherols, leading to the mistaken assumption than tocopherols were only present in photosynthetic parts of plants. The occurrence of tocopherols in roots, nuts, and seeds was further evidenced [
70,
71]. Regarding tocotrienols, they are present in kernels of most monocotyledons, including cereals and absent in the other parts of plants as a result of the specific location of the homogentisate geranygeranyltransferase enzyme in plastid cell of seed endosperms. Data related to the distribution of tochochromanols within cereal kernels are gathered in
Table 2. It clearly appears that tocopherols are primarily found in germs, representing more than 89% of the total tocochromanol content; this percentage reaches 98% when considering germs of maize kernels. Moreover, as indicated in
Table 2, tocotrienols are predominantly present in endosperm (up to 82% of the total tocochromanol concentration) and in pericarp (up to 78%), except for maize kernels, where the pericarp contains mainly tocopherols. The major isomers found in the different parts of kernels differ depending on the considered cereal. In maize, the γ- isomers of both tocopherol and tocotrienol are predominant in all parts of kernels, with a prevalence of γ-tocopherol in the germ and pericarp (65% and 39%, respectively, of total tocochromanol content) and a majority of γ-tocotrienol in the endosperm (45%) [
58,
72,
73].
In wheat germ, the α- isomer of tocopherol is the most representative tocochromanol, reaching up to 68% of the total, followed by β-tocopherol (25%) and β-tocotrienol (6%). In endosperm and pericarp, β-tocotrienol is the most important tocochromanol (49 and 55%, respectively), followed by γ-tocopherol in endosperm (37%) and α-tocotrienol in pericarp (21%) [
54,
73,
74]. In barley, the α-isomers are the predominant isomers in all parts of the kernels: α-tocopherol in germ (68%) and α-tocotrienol in both endosperm and pericarp (41 and 47%) [
73]. In the germ of rice kernels, α-tocopherol is largely predominant, accounting for more than 80% of the total tocochromanol content. In the endosperm of rice kernels, α-tocopherol and γ-tocotrienol are equally represented (39%) followed by α-tocotrienol (21%), while in pericarp α-tocopherol is the most represented tocochromanol (37%) followed by γ-tocotrienol and α-tocotrienol (29 and 27%, respectively) [
73].
5. Kinetics of Tocochromanol Accumulation during Maturation of Cereal Kernels
There are very few dynamic studies that have addressed the tocochromanol composition of kernels from the early stages of kernel development until maturity. This knowledge is, however, essential to clarify which compounds fungal pathogens face at the onset of infection and to support their potential role in plant defense. In maize, several studies corroborate an increase of tocochromanol content starting from pollination. In the report of Xie et al. [
56], focusing on the first 30 days following pollination of maize, a regular increase of both tocopherol and tocotrienol was described. Increase in tocopherol content was shown to continue until 96 days after silking by Picot et al. [
75]. In oat kernels, a pronounced increase in tocochromanols was reported from 14 days after anthesis to the maturity stage [
60]. In barley, tocochromanols were reported to be at their highest concentration at a very early stage (milk stage) to then slightly decrease until maturity [
76]. In rice, the kinetics of tocochromanol content during seed filling were significantly different between tocopherols and tocotrienols; tocopherols were reported to decrease, while an inverse tendency was described for tocotrienols [
66].
This entry is adapted from the peer-reviewed paper 10.3390/ijms23169303