Cancer is the second leading cause of death after cardiovascular pathologies. The incidence of cancer is increasing in both developing and developed countries, and approximately 10 million people died of this deadly disease in 2018 [
1]. The increased prevalence of cancer is often correlated with hereditary traits, environmental pollution, bad lifestyles with poor dietary habits, and a lack of physical activity, leading to the oncogenic transformation of cells [
2]. Epigenetic alterations play a pivotal role in these transformations, which are heritable changes in gene functioning without altering the DNA sequence [
3]. These alterations tend to disrupt the overall balance between tumor-suppressor genes and oncogenes in favor of the latter, thus promoting cellular proliferation and transformation. Because of their reversible nature, scientists are actively investigating the aberrant epigenetic mechanisms in cancer to target them for therapies [
4]. Key epigenetic mechanisms include DNA methylation, histone modifications, and non-coding RNA regulation [
5,
6]. Aberrant DNA methylation and histone modifications play a major role in tumorigenesis and the epigenetic mediators involved in these mechanisms have been well explored for anticancer targeting.
2. Structure and Function of UHRF1
2.1. Structure of UHRF1
Human UHRF1 (initially known as ICBP90) was discovered by two of us as a transcriptional regulator of topoisomerase IIα by binding to an inverted CCAAT box in its promoter region [
10,
11]. However, it is now well-characterized for its involvement in various epigenetic and cellular pathways, including the maintenance of methylation patterns on DNA, histone modifications, DNA damage repair, and the regulation of other proteins [
12,
13,
14]. UHRF1 is a 793-amino-acid-long protein coded by the
UHRF1 gene mapped at the 19p13.3 location in the human genome (
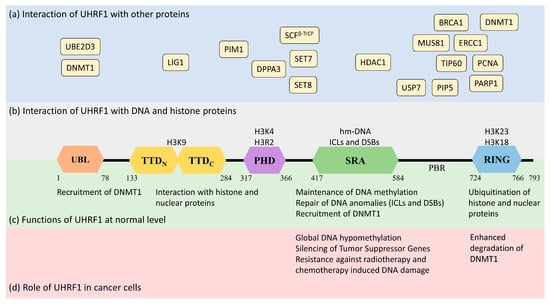
Figure 1). UHRF1 is a protein with multiple domains that differ in their structure and functions [
15,
16]. Starting from the N-terminus, the domains are the ubiquitin-like domain (UBL), tandem Tudor domain (TTD), plant homeodomain (PHD), set and ring-associated (SRA) domain, and, finally, the really interesting new gene (RING) domain at the C-terminus of the protein (
Figure 1).
Figure 1. Structure and functions of UHRF1 protein (isoform 1) in normal and cancer cells along with its interaction with the DNA genome and nuclear proteins. UHRF1 is a multidomain protein having ubiquitin-like domain (UBL), tandem Tudor domain (TTD), plant homeodomain (PHD), SET and RING-associated domain (SRA), and really interesting new gene domain (RING) in its structure. (a) (in blue) indicates the interaction of different nuclear proteins with the proposed domains of UHRF1. (b) (in grey) indicates the interaction of UHRF1 with DNA and histone proteins. The SRA domain of UHRF1 recognizes hemi-methylated DNA or anomalies in DNA structure (ICLs and DSBs). TTD, PHD, and RING domains of UHRF1 interact with the indicated amino acids in H3 histone proteins. (c) (in green) highlights the cellular functions of UHRF1 in normal cells. UHRF1 is implicated in DNA methylation maintenance by recruiting DNMT1, recognizing DNA damage and initiating the DNA damage response, and regulating the function and stability of nuclear proteins through ubiquitination. (d) (in red) indicates the role of UHRF1 in cancer cells. UHRF1 represses many tumor-suppressor genes by maintaining the hyper-methylation of their promoters. High levels of UHRF1 in cancer cells also promote genetic instability by destabilizing DNMT1, which induces global hypomethylation. Furthermore, increased levels of UHRF1 facilitate the repair of DNA damage induced by radio or chemotherapy, making cancer cells resistant to anticancer therapy.
It is interesting to note that UHRF1 shares a full sequence identity of 52% with UHRF2, another member of the UHRF family of proteins. The sequence identity is highest between the RING (79%) and the SRA (77%) domains of UHRF1 and UHRF2. However, despite their high sequence similarity, the expression profiles, the distributions, and the functions of these proteins are different [
17].
2.1.1. Ubiquitin-like Domain (UBL)
The UBL is structurally 35% identical to ubiquitin and is linked to the stability of UHRF1. It possesses characteristic α-helix and β-sheets, as well as two conserved lysine residues, K31 and K50 (similar to ubiquitin K29 and K48), which can be polyubiquitinated to trigger proteasomal degradation [
12]. A role of the UBL has also been identified in DNMT1 (DNA methyltransferase 1) recruitment and the accurate transmission of methylation patterns [
16]. Indeed, the UBL coordinates with the UHRF1 RING domain to ubiquitinate histone H3 residues, which serve as an anchorage signal for DNMT1 recruitment on hemi-methylated DNA [
18,
19]. Interestingly, the direct interaction of UHRF1-UBL with the RFTS (Replication Focus-Targeting Sequence) domain of DNMT1 also facilitates the enzymatic activity of the latter by activating its catalytic domain [
20].
2.1.2. Tandem Tudor Domain (TTD)
The Tandem Tudor domain of UHRF1 is involved in various protein–protein interactions vital for UHRF1 biological functions. The TTD notably allows UHRF1 to interact with the histone marks needed for its functioning [
21]. The TTD is made up of two subdomains (TTDN and TTDC), each having a characteristic five-stranded β-barrel moiety in its structure [
22]. The aromatic cage (Phe-152, Tyr-188, and Tyr-191) in TTDN, along with Asn-194 and Asp-145, recognizes di- and tri-methylated H3K9, while the peptide-binding groove between the TTDN and TTDC is sensitive to the epigenetic modifications on adjacent histone residues (i.e., H3K4 methylation and H3T6 phosphorylation) [
22]. Through its interaction with LIG1 (DNA ligase), the TTD allows UHRF1 recruitment to replication foci to play its role in DNA methylation [
23], but this interaction is not essential for abnormal DNA methylation patterns in cancerous cells [
24]. The TTD also interacts with LIG1K126me3, which opens the closed conformation of UHRF1 [
25,
26]. In contrast, when the TTD interacts with the polybasic region (PBR) of UHRF1, the binding of H3K9me3 with the TTD-PHD domains is disrupted, ultimately returning to the closed conformation of UHRF1. Finally, when UHRF1 binds to hemi-methylated DNA or USP7 (Ubiquitin-Specific-processing Protease 7), the interaction of the TTD with the PBR is disturbed, which favors the transition to the open conformation of UHRF1 [
27,
28,
29].
2.1.3. Plant Homeodomain (PHD)
The plant homeodomain (PHD) of UHRF1 differs from the canonical PHDs in other proteins by having a PHD motif containing four cysteines (C302, C305, C313, and C316) in a loop coordinated with a zinc atom linked to the canonical PHD region by a single helical turn. The PHD domain of UHRF1 specifically recognizes the H3R2 motif in chromatin, which is essential for UHRF1 functions [
30,
31]. This interaction of the PHD with histone proteins can be altered through other proteins, such as DPPA3 (developmental pluripotency-associated protein 3), inhibiting UHRF1 localization on chromatin and promoting passive demethylation [
32].
2.1.4. Set and Ring-Associated (SRA) Domain
The SRA is a highly conserved domain specific to the UHRF family of proteins [
12], playing an important role in DNA methylation [
33,
34,
35]. Through this domain, UHRF1 recognizes hemi-methylated DNA and flips the methylated cytosine out of the helix. The SRA domain functions as a palm of a hand grasping the DNA duplex, where the NKR finger (489–491 amino acid residues) and thumb (444–496 amino acid residues) form two specific loops that project into major and minor grooves of the DNA double helix to read the nucleotides in the CpG duplex. The NKR finger specifically identifies the hemi-methylated DNA and flips the methylated cytosine out of the duplex. The flipped methylated cytosine is later stabilized by π-π stacking with the conserved tyrosine (466 and 478) residues in the binding domain [
33]. The NKR domain also helps UHRF1 to differentiate between hemi-methylated and fully methylated DNA, since the second methylated cytosine creates a steric hindrance for the NKR finger, leading to a reduced affinity of UHRF1 for methylated DNA [
33]. The only other protein from vertebrates carrying an SRA domain is UHRF2 [
11]. In contrast to UHRF1, UHRF2 exhibits tumor-suppressor gene capacities [
11,
17], but an ability to favor tumor progression cannot be excluded, at least in certain types of cancer, such as hepatocellular carcinoma [
36] or intestinal tumorigenesis [
37]. The homology of amino-acid sequences between UHRF1 and UHRF2 reaches 74% [
11,
17], questioning the possibility that a drug targeting the SRA domain of UHRF1 putatively also may bind to the SRA domain of UHRF2 and thus may have opposite pharmacological effects. This seems, however, unlikely, but not impossible, considering the structures of each SRA domain. First, the UHRF2 SRA domain shows preferential recognition of hydroxymethyl cytosine over methylcytosine [
38], suggesting differences in the respective structures. Indeed, the NKR loop, involved in the base-flipping mechanism, is disordered in the UHRF2-SRA domain, while in the UHRF1-SRA domain, it is not. This difference has been proposed to explain why the UHRF2-SRA domain has a preference for fully hydroxymethylated DNA over hemi-hydroxymethylated DNA vs. the UHRF1-SRA domain having a preference for hemi-methylated DNA over fully methylated DNA [
38]. Altogether, this supports the fact that drugs targeting the SRA domains of UHRF1 would have specificity versus UHRF2, despite their strong similarities (77% amino acid sequence identity, personal observations).
2.1.5. RING Domain
The RING domain harbors the only enzymatic activity of this protein [
39]. It is rich in cysteine residues that form two zinc fingers interacting with a variety of substrates. Through this domain, UHRF1 can ubiquitinate itself, but also DNMT1, H3, and other proteins, regulating their function and stability [
19,
40].
2.2. Functions of UHRF1
UHRF1 is an important component of an epigenetic complex acting after DNA replication. UHRF1 is primarily involved in the maintenance of genomic DNA methylation patterns in cells [
12,
41]. Via its SRA domain, UHRF1 recognizes the CpG motifs on the parent strand of hemi-methylated DNA and flips the methylated cytosine out of the duplex. This then enables DNMT1 to methylate the opposite cytosine on the newly formed daughter strand [
33,
34,
35,
42]. UHRF1 also helps in recruiting DNMT1 to the hemi-methylation sites by direct interaction through its UBL and SRA domains, or indirectly by ubiquitinating histone H3K18 through its RING domain, as H3K18ub serves as a binding site for DNMT1. The crosstalk of UHRF1 TTD with H3K9me3 and H3K4 or PHD with H3R2 also plays a significant role in the recognition of the methylation site and the maintenance of the methylation patterns on the daughter strand [
22,
43,
44,
45]. The recruitment and activity of UHRF1 at the replication site is also facilitated by the DNA replication machinery. DNA ligase 1 (LIG1), methylated by G9a and GLP methyltransferases, mimics H3K9me2/3 in binding to the TTD of UHRF1. This binding recruits UHRF1 to the DNA replication sites for the maintenance of DNA methylation [
23]. Similarly, the interaction with hemi-methylated DNA, USP7, and PIP5 (Phosphatidylinositol-4-phosphate 5) during the S-phase, triggers UHRF1 to switch from its “closed” to its “open” state, which facilitates UHRF1 loading onto the newly formed DNA [
27,
29,
46] (
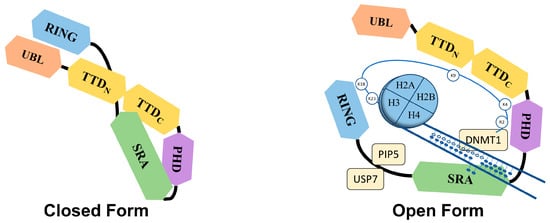
Figure 2).
Figure 2. Proposed closed and open conformations of UHRF1. The PBR region between the SRA and RING domains interacts with TTD, which prevents the interaction of TTD with H3K9me3 marks, thus keeping the UHRF1 in closed conformation. On the other hand, proteins like PIP5 and USP7 can interact with the PBR of UHRF1 and disrupt the association between PBR and TTD, allowing TTD to interact with H3K9 methylation marks, which renders the UHRF1 in an open conformation. This open conformation of UHRF1 is also promoted by the binding of the SRA domain with hemi-methylated DNA, particularly in the S phase of the cell cycle.
Along with its role in epigenetics, UHRF1 is also implicated in the DNA damage response and helps to maintain the integrity and stability of the genome. Initially, it was observed that the inhibition or depletion of UHRF1 leads to increased sensitivity to irradiation and higher accumulation of γH2AX in irradiated cells [
47,
48]. This function of UHRF1 in the DNA damage response is attributed to its SRA and RING domains, where the SRA domain acts as a sensor for DNA damage and the RING domain ubiquitinates the interacting proteins and directs the DNA repair pathways [
47,
48].
Owing to its SRA domain, UHRF1 identifies interstrand crosslinks (ICLs) and promotes DNA damage repair [
49,
50]. UHRF1 accumulation at damage sites precedes the recruitment of important effector proteins, such as FANCD2 (Fanconi anemia complementation group D2), which, in turn, is necessary for the recruitment of other factors of the Fanconi anemia (FA) repair pathway [
50]. UHRF1, along with its paralogue UHRF2, forms a complex that monoubiquitinates FANCD2, which further enhances its retention after recruitment at the site of DNA damage [
51]. UHRF1 also aids in the recruitment of ERCC1 (excision repair cross-complementation group 1) and MUS81 (crossover junction endonuclease MUS81 enzyme) at the site of ICLs through direct interactions with the SRA and RING domains, working as a nuclease scaffold to facilitate the ICL repair independently of the FA pathway [
49]. It is very important to note that the recognition of ICLs by UHRF1 and facilitation in the DNA damage response is more pronounced in the S phase of the cell cycle and with ICLs showing minor distortions.
UHRF1 is also associated with DNA damage repair resulting from double-strand breaks (DSBs), where it facilitates DNA repair by homologous recombination (HR) [
52]. During DNA replication, UHRF1 accumulates at DSBs through its interaction with the BRCT (BRCA1 C-terminal) domain of BRCA1 (breast cancer gene 1) and phosphorylation of UHRF1 at Ser 674 by CDK2/cyclin. UHRF1, in turn, ubiquitinates the replication timing regulatory factor 1 (RIF1) at lysine K63, which results in a disruption of RIF1/53BP1 (p53-binding protein 1) foci formation, favoring DNA damage repair through the homologous recombination (HR) pathway by the accumulation of BRCA1 protein at the site of damage. Later, it was also reported that phosphorylated UHRF1 was prone to methylation by SET7 (histone-lysine N-methyltransferase) during the S phase, which enhanced the interaction between UHRF1 and PCNA (proliferating cell nuclear antigen) proteins, resulting in polyubiquitination of PCNA, thus stimulating DNA repair by the homologous recombination pathway [
53]. This presence of DNA damage repair through the HR pathway is further augmented by the interaction of methylated UHRF1 with PARP1 [(poly(ADP-ribose) polymerase 1)], promoting cell cycle progression and increasing the DNA repair efficiency [
54]. From all these studies, it can be concluded that UHRF1 undoubtedly plays an important role in DNA repair, but the intimate mechanism in which it is involved remains a mystery. Does it play a role in DNA sequence repair and/or the restoration of the DNA methylation patterns on the repaired DNA fragments? Further studies are required to fully decipher its role in DNA repair processes.