1. Introduction
More and more nanomaterials have been developed and intended for in vivo applications, such as disease diagnosis and treatment with improved sensitivity and efficiency in biomedicine, taking advantage of the unique physical and chemical properties of nanomaterials [
1,
2]. At the same time, even more nanomaterials not intended for in vivo applications may also eventually enter human bodies, and thus the safety assessment of nanomaterials has long been an important issue. Both biomedical applications and safety assessments of nanomaterials require a deep understanding of their behavior in organisms. The first and fundamental step is to understand how nanoparticles (NPs) enter cells, translocate in cells, and leave cells.
The uptake and trafficking of various NPs in cells have been studied extensively and have gained a deep understanding, including the aspects of intracellular uptake, endocytosis pathways, organelle localization, and related influencing factors [
3,
4,
5]. Compared with the investigations on endocytosis, less attention has been paid to the exocytosis of NPs, and sometimes the results obtained are inconsistent in many aspects [
3,
6,
7]. Current exocytosis studies focus on a few NPs such as SiO
2 NPs [
8,
9,
10], Au NPs [
11,
12], quantum dots (QDs) [
13,
14,
15], and polymers [
16,
17]. Unfortunately, most of the time, the exocytosis of NPs is simply measured as a supplement to the cellular uptake and transport of NPs in cells [
18], or even worse, some researchers believe that the exocytosis of NPs is trivial or negligible [
19].
In fact, exocytosis is as important as endocytosis in the interaction of nanomaterials and cells [
17,
20]. It is the process of discharging unwanted cargo and other macromolecules from cells [
21,
22], a process opposite to the endocytosis of NPs [
23]. Understanding the exocytosis of NPs means to answer how and how fast intracellular NPs move out of cells and to what extent intracellular NPs can still remain in the cells. The endocytosis and exocytosis of NPs jointly decide the content of NPs in cells, while the retention of NPs in cells is indirectly and directly related to their cytotoxicity [
11,
24,
25,
26,
27]. NPs trapped in cells for a long period may induce various adverse effects, such as cell growth inhibition, asymmetric intercellular metastasis, and cytotoxicity [
6,
28]. Exocytosis is also related to the translocation of NPs at the animal level. In nanomedicine, researchers design NPs as carriers to deliver therapeutic and imaging agents with the ultimate goal of achieving high-efficacy and high-sensitivity imaging [
29]. To achieve this goal, NPs have to penetrate tissues to reach targets, and the retention of NPs in target cells is critical, as the discharge of NPs that have been engulfed ultimately affects the number of drugs at the site of action [
28,
30,
31,
32]. The longer the intracellular persistence time in target cells, the higher therapeutic potential of the NP-based drug carriers. It is therefore necessary to enhance the retention of NPs in cancerous cells and reduce their retention in healthy cells [
33]. Yet the fate of these NP carriers (after releasing drugs) in cells is largely overlooked. There is a possibility that these NP carriers could be trapped in cells for a long time [
22,
28,
34], and thereby induce cytotoxicity [
16]. Therefore, understanding the process and mechanisms involved in the exocytosis of NPs is necessary and urgent [
35].
2. Exocytosis Mechanisms/Pathways of NPs
Exocytosis is the transport of nonessential molecules from the interior of a cell to the exterior to reduce the stress inside the cell, a process opposite to endocytosis. After NPs enter cells, they may reside and transport in various vesicles and organelles, including the endosome, lysosome, mitochondrion, ER/Golgi apparatus, nucleus, and cytoplasm [
10,
36,
62,
83,
84]. Some NPs remain inside the cell for a long time without exocytosis [
25,
85], which is usually associated with cytotoxicity.
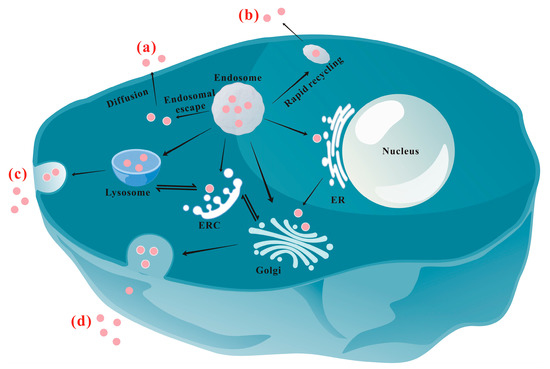
Figure 1 schematically shows the main exocytosis mechanisms/pathways of NPs. At present, the recognized exocytosis pathways of NPs mainly include the following: (a) Diffusion: As an energy-independent pathway, diffusion allows NPs to cross the phospholipid membrane directly and passively [
8,
46,
86,
87]. Fundamentally, diffusion is driven by the concentration gradient of NPs between the inside and outside environment, and the coefficient of diffusion is inversely proportional to the diameter of the NPs. So, generally, only particles smaller than 50 nm have the possibility of entering and exiting the cell through diffusion [
88]. NPs in the cytoplasm may be excreted via diffusion more slowly compared with vesicle-coated NPs [
6,
9,
89], which are much easier to move within cells. The results are similar to those reported by Yang et al. [
90], who found that more than 10 h was needed for the functionalized PS that first distributed in the cytoplasm and then moved to lysosomes to be eliminated by cells. (b) Rapid recycling pathway: Some NPs-containing early endosomes near the cell membrane may directly fuse with the membrane to rapidly release the NPs to the extracellular environment [
44,
91]. Researchers could compel NPs to exocytosis directly from the endosomes by inhibiting the transport of NPs to the perinuclear [
53]. (c) Lysosomal pathway: This is the most important intracellular circulation pathway to release NPs to the extracellular environment [
10,
55]. Actually, both the rapid recycling and the lysosomal routes involve cell membrane fusion [
92]. (d) ER/Golgi apparatus pathway: NPs delivered to the ER or Golgi apparatus are released by secreting vesicles that fuse with the cell membrane [
93,
94,
95]. This is a relatively slow pathway [
7,
83,
96,
97]. (e) Other pathways: Special receptors on the surface of the cell membrane, such as P-glycoprotein and αvβ3 integrin, may interact with NPs to trigger exocytosis through rapid recycling of the receptors [
71,
98]. A survey of 70 available research papers on the exocytosis pathways of NPs reveals that the lysosomal pathway is the most important exocytosis mechanism of NPs (involving 46 papers). The second most important pathway is the ER/Golgi pathway, which is reported in 22 papers. Diffusion also holds a place, as reported in seven papers, while four papers reported different pathways from the above. Certainly, NPs may be excreted through more than one pathway, which was observed in 11 papers.
Figure 1. Schematic illustration of the main exocytosis mechanisms/pathways of NPs: (a) Diffusion; (b) Rapid recycling pathway; (c) Lysosomal pathway; (d) ER/Golgi apparatus pathway. ERC: endocytic recycling compartment; ER: endoplasmic reticulum.
In addition, it is worth noting tubulin plays an important role in the exocytosis of NPs. In fact, the microtubule acts as a kind of “courier” that is involved in the whole process of exocytosis [
42,
58,
99].
2.1. Lysosomal Pathway
The lysosome is the terminal degradative compartment of the endocytic pathway. After internalization, most NPs will be eventually trapped in lysosomes [
9,
10,
15]. As shown in
Figure 1, most of the NP-containing endosomes will mature into lysosomes, where some NPs might be degraded. If not degraded, cargoes in the lysosomal system are expected to be recycled back to the cell surface [
45]. Stable NPs in lysosomes would either enter the Golgi apparatus via the endocytic recycling compartment (ERC) for excretion or undergo lysosomal exocytosis directly [
100]. It has also been reported that inhibiting the autophagy could prevent the accumulation of polystyrene (PS) NPs in lysosomes and thereby egress in live alveolar epithelial cells [
55], indicating NPs in autophagic vacuoles could be excreted via the lysosomal pathway after autophagic vacuoles fuse with lysosomes [
66,
101].
The vesicle-packed NPs are excreted from the lysosome when the vesicle approaches and fuses with the cell membrane [
10]. For example, Yanes et al. found that the efflux of mesoporous SiO
2 NPs (MSNs) in several types of cells was mainly through lysosomal exocytosis [
10]. Phosphonate-modified MSNs (P-MSNs) in the cells dramatically decreased to less than 40% of the control after 6 h in the P-MSN-free medium, and almost all intracellular MSNs were excreted after 48 h. The exocytosis in different cells could be inhibited by nocodazole (
Table 1, inhibit the transport of the lysosome to the periphery and fusion with the plasma membrane) and the excretion of P-MSNs positively correlated to that of the lysosome secretion marker β-hexosaminidase, indicating it is a lysosomal exocytosis process. Similarly, Chu et al. systematically studied the excretion of 50 nm SiO
2 NPs by preincubating cells with NPs for 48 h and then incubating them in an NP-free medium for 1 h [
9]. TEM investigation indicated that, after 48 h incubation, the exocytosis of NPs occurred along with endocytosis, and most NPs accumulated in lysosomes. When the NP-laden cells were cultured for 1 h in an NP- and serum-free medium, the number of NPs in lysosomes decreased dramatically, suggesting the excretion of NPs through the lysosomal pathway. The TEM result was confirmed by the decrease in the fluorescence intensity of NPs in cells measured using CLSM. The exocytosis of functionalized MSNs in 4T1 cells was also mediated by the lysosomal pathway rather than the Golgi apparatus pathway because Brefeldin A (
Table 1, causes the collapse of the Golgi apparatus) did not inhibit the exocytosis of MSNs [
102]. Liu et al. reported that the exocytosis of 50 nm PS and 500 nm PS were decreased to 33% and 40% of the control group, respectively, by the inhibitor Bafilomycin A1, while the exocytosis was promoted to 125% (50 nm PS) and 148% (500 nm PS) by the accelerator ionomycin [
103], confirming the lysosomal exocytosis of these PSs. Similarly, chemical inhibition studies have confirmed that the lysosomal pathway is the main exocytosis pathway for polyethylene glycol (PEG)-phospholipid-coated upconversion NPs in HeLa cells [
42] and polyamidoamine (PAMAM-NH
2) NPs in MCF-7/ADR cells [
56].
It should be noted that the decrease in NPs in lysosomes cannot be directly attributed to the exocytosis of NPs via the lysosomal pathway. NPs in lysosomes may be delivered to other organelles, such as the Golgi apparatus (
Figure 1) [
37,
44,
83]. In addition, NPs in lysosomes may undergo degradation (such as the detachment of surface modification groups) and then can hardly be involved in exocytosis or intracellular trafficking, resulting in a longer intracellular retention [
32,
104].
Table 1. Commonly used concentrations and mechanisms of exocytosis inhibitors and accelerators.
| Chemical |
Concentration |
Pathway |
Function |
Ref. |
| Inhibitor |
NaN3/4 °C |
-- |
-- |
Inhibiting cell membrane fluidity and energy-dependent transport |
[54] |
| Bafilomycin A1 |
0.1~0.5 μM |
Lysosomal |
Inhibitor of endosomal acidification and lysosomal maturation |
[55,56] |
| Chloroquine |
100 μM |
Lysosomal |
pH buffering and inhibiting lysosomal enzymes |
[57] |
| LY294002 |
250 nM~1 mM |
Lysosomal |
Inhibiting PI3 kinase and lysosomal exocytosis |
[10,37] |
| Vacuolin-1 |
5 μM |
Lysosomal |
Inhibiting Ca2+-dependent lysosomal exocytosis |
[37] |
Methyl-β-cyclodextrin
(MβCD) |
1~10 mM |
Lysosomal |
Cholesterol depletion |
[58,59] |
| Wortmannin |
10~33 µM |
Lysosomal |
Preventing the transport of endosomes to lysosomes |
[17,60] |
| U1866A |
2.5 µM |
Lysosomal |
Altering cholesterol accumulation and affecting different intracellular trafficking pathways |
[10] |
| Monensin |
14~50 μM |
Golgi |
Blocking transportation from the Golgi apparatus to the cell membrane |
[56,61] |
| Exo1 (2-(4-fluorobenzoylamino)-benzoic acid methyl ester) |
10~100 µM |
Golgi |
Inducing collapse of the Golgi apparatus |
[10,62] |
| Brefeldin A |
35~90 μM |
ER/Golgi |
Inhibiting transport from the ER to the Golgi apparatus |
[42,56] |
| Nocodazole |
15~33 μM |
microtubule-associated transport |
Inhibiting microtubule formation |
[37,59] |
| Cytochalasin D |
5 µM |
-- |
Disruption of actin polymerization |
[37] |
| Verapamil |
10 µM~2 mM |
-- |
P-glycoprotein inhibitor |
[56,63] |
| Fumitremorgin C |
5 µM |
-- |
MDR-associated protein inhibitor |
[56] |
| Accelerator |
Ionomycin |
10 μM |
Lysosomal |
Accelerating exocytosis by transporting Ca2+ into the cells |
[64] |
| A23817 |
-- |
Lysosomal |
Promoting Ca2+ influx and accelerating lysosomal associated efflux |
[65] |
2.2. ER/Golgi Pathway
In addition to lysosomal exocytosis, the secretory pathway containing the Golgi apparatus (and ER) is another important exocytosis route [
32,
105]. In this pathway, cells eject ingested contents through certain specific secretory vesicles, which is also believed to be a safe route for drug delivery that escapes the degradation of lysosomes [
106]. NPs transported from the ER or other different places such as lysosomes first arrive at the Golgi apparatus and then are transported to the cell membrane. In the literature, some studies may only report that NPs are excreted via the Golgi apparatus, without analyzing whether they pass through the ER. For example, by using vesicle marker GTPases to investigate the colocalization of WS
2 nanosheets (NSs) with different vesicles, Kong et al. found that WS
2 NSs in early and late endosomes (Rab22- and Rab24-labeled, respectively) might be delivered to the Golgi, and then the WS
2 NSs could be excreted into the extracellular space with the help of classic (Rab26- and Rab3-labeled) and GLUT4 (Rab10- and Rab8-labeled) translocated vesicles [
83]. The excretion mechanism was confirmed by inhibiting the exocytosis using the exocytosis inhibitor Exo1, which induces the collapse of the Golgi apparatus (
Table 1). Similarly, Ding et al. reported that Exo1 inhibited the excretion of intracellular fluorescent MSNs, and the fluorescent MSNs merged with Rab3 and Rab26 (marking the transfer from the Golgi to the cell membrane) marked secretory vesicles well shown by CLSM, indicating the involvement of the Golgi exocytosis pathway in their efflux [
62].
Whether NPs pass through the ER to the Golgi and then are expelled can be identified by using specific chemical inhibitors or trafficking vesicle markers (such as Rab1, Rab2, and Rab43) [
56,
66]. For example, Liu et al. found that the discharge of intracellular CdSe/ZnS QDs functionalized with 2-mercaptoethylamine (MEA-QDs) could be dramatically suppressed when Brefeldin A and Monensin were used to block the ER and Golgi pathways, respectively, indicating that both the ER and Golgi apparatus are crucial regulatory stations in the exocytosis of MEA-QDs [
37]. Xing et al. observed that the inhibitor Brefeldin A could block the exocytosis process of L-cysteine (Cys) and cell-penetrating peptide octa-arginine (R8)-modified insulin-loaded NPs, suggesting that the NPs were excreted via the pathway from the ER/Golgi apparatus to the cell surface [
99]. Further studies indicated that Cys modification was critical for the exocytosis through the Golgi secretory pathway due to the thiol group of Cys binding to the sulfhydryl receptor site in the Golgi apparatus.
The golgi apparatus is an important regulator which participates in the ER/Golgi apparatus pathway and Golgi apparatus/plasma membrane pathway. During the process of exocytosis, the ER and Golgi apparatus secrete some NP-containing vesicles, while the vesicle-SNARE (v-SNARE) protein interacts with the target-SNARE (t-SNARE) protein in the plasma membrane, followed by further membrane fusion [
107,
108]. As the proteins widely spread in the plasma membrane–Golgi–ER recycling pathway, SNARE proteins are responsible for membrane fusion [
32]. Lysosomal exocytosis also involves a similar process and corresponding SNARE proteins [
109].
2.3. The Auxiliary Effect of the Microtubule
Except for passive diffusion, the microtubule is involved in all the other exocytosis pathways of NPs. During exocytosis, NPs in endosomes, lysosomes, the ER/Golgi, or other compartments move out of the cell with the help of microtubules [
13,
42,
62]. In fact, ERC is a microtubular organelle distributed in the cytoplasm, which is an important chamber that takes part in the formation, fusion, and release of vesicles. NPs transported to the ERC can be recycled back to the plasma membrane via recycling vesicles and/or be directed to lysosomes, the ER/Golgi apparatus, and other organelles.
In general, NPs encapsulated in vesicle systems are delivered from the cell periphery to the perinuclear region by dyneins along microtubules, while their delivery from the perinuclear region to the cell periphery is operated by kinesins along microtubules [
110,
111]. In the excretion process, NPs accumulated in microtubules near the nucleus gradually disperse from the accumulation site and then travel along microtubules to nearby cell membranes [
112], where they are expelled [
42]. For example, Cao et al. reported that most of Si-NPs located in early endosomes were transported to lysosomes and the ER/Golgi with the aid of microtubules [
67]. Liu et al. also observed that the exocytosis of carbon dots (CDs) in five types of cells was microtubule-dependent [
37], since Nocodazole significantly inhibited the efflux of CDs from the cells. Malik et al. studied the trafficking and exocytosis of PNA (peptide nucleic acids, 180 nm) in HeLa cells using CLSM and flow cytometry and found that the interruption of the recycling pathway of PNAs via inhibiting either Rab11a or Rab27b directed the intracellular trafficking toward the lysosomal stage, while Rab11a and Rab27b were associated with microtubule organelle ERC [
44].
The pathways chosen by a cell to eliminate NPs are influenced by a variety of factors, including the physiochemical properties of NPs, cell type, extracellular conditions, etc., which will be discussed later in this research Currently, there is no widely recognized conclusion about which parameters regulate the intracellular NPs to leave cells via specific pathways. Basically, the uptake pathways of NPs decide their exocytosis in a specific cell line. For example, NPs entering cells via caveolin-mediated endocytosis mainly enter the ER, and then most of them leave the cells via the ER/Golgi apparatus. However, some of them may also leave the ER and enter lysosomes, following exocytosis via lysosomal exocytosis. Exocytosis is complex and the related mechanisms need further in-depth research.
It is worth noting that exocytosis experiments are generally performed under nontoxic conditions to eliminate the influence induced by cell damage [
37]. The cytotoxicity of NPs has potential effects on exocytosis, for example, people can reasonably speculate that damage to the cell membrane may accelerate the exocytosis of NPs. Gupta et al. [
113] reported that nanosized extracellular vesicles were excreted from normal cells through Rab protein-dependent lysosomal exocytosis, but those vesicles were excreted from necroptotic cells through the mixed-lineage kinase-domain-like (MLKL)-mediated calcium influx induced lysosomal exocytosis (Rab protein-independent), indicating the status of cells affects the exocytosis process. However, as cytotoxicity is complex and difficult to be precisely regulated, exocytosis studies are performed generally on cells without an obvious viability loss, though it is interesting to investigate the effect of cytotoxicity on the exocytosis of NPs.
This entry is adapted from the peer-reviewed paper 10.3390/nano13152215