Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
LOXL2, a copper-dependent amine oxidase, has emerged as a promising therapeutic target in hepatocellular carcinoma (HCC). Increased LOXL2 expression in HCC has been linked with an aggressive phenotype and represents a poor prognostic factor.
- hepatocellular carcinoma (HCC)
- LOXL2
- tumor microenvironment (TME)
- extracellular matrix (ECM)
1. Introduction
Hepatocellular carcinoma (HCC) is the most common type of primary liver cancer, accounting for more than 80% of cases [1]. With incidence and mortality rates increasing worldwide, it represents a global healthcare concern [1,2]. The underlying process in the development of HCC is chronic liver damage leading to cirrhosis, which is most often caused by chronic viral hepatitis (hepatitis B and C), alcohol abuse, and metabolic dysfunction-associated steatotic liver disease (MASLD) [3]. Around 80% of HCC arises from a cirrhotic liver, and among patients with cirrhosis of any etiology, one-third will develop HCC [1,4,5]. The pathogenesis of HCC is a complex multistep process that usually starts with liver injury and inflammation. Chronic liver inflammation triggers a fibrous process, which over time can progress to cirrhosis and is characterized by the extensive disruption of liver tissue architecture. Prolonged inflammation and cirrhosis create precancerous settings resulting in the generation of dysplastic foci and dysplastic nodules, which further accumulate genetic/epigenetic alterations, resulting in the development of hepatocellular carcinoma [6].
The most appropriate therapeutic option is determined based on the TNM tumor stage, degree of background liver damage, and the patient’s overall health [7]. The recommended therapy for an early-stage HCC is surgical resection, liver transplantation, or radiofrequency ablation. Transarterial chemoembolization (TACE) and radiation therapy, alone or in combination with systemic therapy, are used for intermediate-stage HCCs, while systemic therapy (chemotherapy, molecular targeted therapy, immunotherapy, and gene therapy) is the treatment of choice for advanced-stage HCCs [7,8]. Unfortunately, due to the asymptomatic nature of early-stage HCC, more than 60% of cases are detected in the intermediate or advanced stage, where curative therapeutic options are limited [9]. The backbone of systemic therapy for HCC is sorafenib and other multi-kinase inhibitors [7,10,11,12]. Moreover, ramucirumab, a vascular endothelial growth factor (VEGF) inhibitor and immune checkpoint inhibitors (nivolumab and pembrolizumab) are being increasingly used, either separately or in combination [7,13]. Although the choices for therapeutic strategies in advanced HCC are rapidly expanding, they provide a variable but still limited extension of survival, cause a wide range of side effects, and ultimately, lead to the development of tumor resistance, which is recognized as one of the biggest problems in the treatment of HCC [10].
The multitude of underlying mechanisms that are responsible for the development of resistance to therapy by HCC is not completely understood, although it has been shown that tumor microenvironment (TME) plays an important role. The tumor microenvironment has a crucial role in hepatocarcinogenesis and directly participates in the regulation of liver fibrosis and tumor-progressive processes, such as epithelial–mesenchymal transition (EMT), extracellular matrix (ECM) remodeling, migration, invasion, and metastasis [14]. Unraveling the complex interactions within the tumor microenvironment and targeting the components of the TME, such as ECM remodeling enzymes, might serve as a valuable strategy to improve the current therapeutic options and develop novel ones while attempting to re-sensitize resistant tumors to existing therapeutic agents [10,15,16].
In the last fifteen years, lysyl oxidase-like 2 (LOXL2) has emerged as one of the major mediators between tumor cells and TME. This research has implicated the involvement of LOXL2 in every step of tumor progression [17,18,19,20,21,22]. The involvement of LOXL2 has been reported in the regulation of cancer cell proliferation, epithelial–mesenchymal transition, migration, extravasation, and creating premetastatic niches at distant sites, as reviewed by Zhang et al. [23], Lin et al. [24], and Wen et al. [25]. The mechanisms through which LOXL2 affects tumor invasiveness can be used as a typical model of solid cancer progression and spreading [26]. In addition, LOXL2 regulates tumor-induced angiogenesis [27,28,29] and mediates the interaction between cancer cells and cancer-associated fibroblasts (CAF), and macrophages [22,30,31]. Taken together, LOXL2 represents a multifunctional protein, which is enrolled in almost every step of solid tumor propagation.
2. LOXL2 Introduction: LOX Family, Structure, and LOXL2 Function
LOXL2 is a secreted and copper-dependent amine oxidase that belongs to the lysyl oxidase (LOX) family, which consists of five homologous members: LOX and LOX-like l–4 (LOXL1–4) proteins [32,33,34,35,36,37]. The primary function of the LOX family enzymes is to catalyze the cross-linking of elastin and collagen by oxidation, which is essential for maintaining the rigidity, stability, and remodeling of the extracellular matrix (ECM) [38,39]. The human LOXL2 gene is positioned on chromosome 8p21-22 and encodes a 774 amino acid protein [26,40]. Structurally, the LOXL2 protein contains a variable N-terminal region and a highly conserved C-terminal region with catalytic activity (Figure 1). The catalytic domains at the C-terminus are conserved among LOX family members and consist of the copper-binding domain, lysyl tyrosine quinone (LTQ) element, which is required for cofactor formation, and a cytokine receptor-like (CRL) domain [41,42]. The LOXL2 N-terminal domain is more variable and includes a signal peptide and four scavenger receptor cysteine-rich (SRCR) elements, which are responsible for protein–protein interactions [41,43].
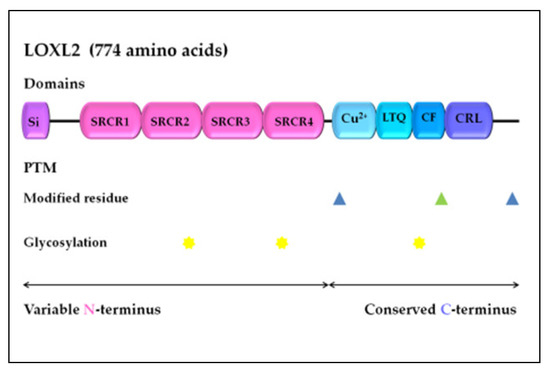
Figure 1. LOXL2 protein structure. Arrows indicate the variable N-terminal domain and conserved C-terminal domain consists of several different functional domains, represented by boxes. Post-translational modifications of LOXL2 were obtained using UniProt. Modified residues are represented by blue triangles (phosphoserine at 601 and 722) and green triangle (2′,4′,5′-topaquinone at 689), while N-linked glycans at N288, N455, and N644 are marked with yellow stars. Si, signal peptide; SRCR, scavenger receptor cysteine-rich domain; Cu2+, copper binding domain; LTQ, lysine tyrosylquinone cofactor residues; CF, cofactor formation; CRL, cytokine receptor-like domain; PTM, post-translational modifications.
The existence of several different protein domains in the LOX family of enzymes implies their multiple biological functions [26], in addition to maintaining the structure of the ECM. The LOX and LOXL1–4 proteins are enrolled in other biological processes, including embryogenesis and development [42,45,46]. Although LOXL2 is a secretory protein, it is also distributed in the intracellular compartments and within the nucleus, thereby can exert its intracellular and extracellular activities through various downstream pathways (Figure 2a) [25]. LOXL2 participates in the structural maintenance of the ECM in fibrotic tissues, such as in the liver tissue [47,48,49]. LOXL2-specific tissue expression among various liver cell compartments can be estimated using multi-omics datasets, as presented in Guilliams et al. [50], which detects the overall expression of LOXL2 in 0.7% of healthy adult liver cells, dominantly in stromal and endothelial cells, as well as in hepatocytes and cholangiocytes (Figure 2b) [51].
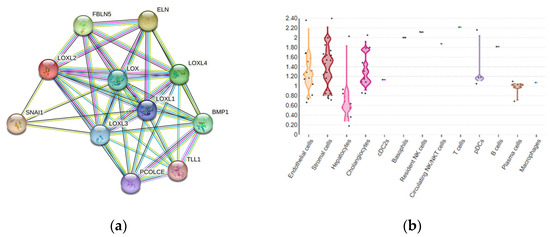
Figure 2. (a) LOXL2 protein interactome. Available on the String platform. SNAI1, zinc finger protein involved in the induction of the epithelial–mesenchymal transition (EMT); ELN, elastin; FBLN5, fibulin-5, which is essential for elastic fiber formation; PCOLCE, procollagen C-endopeptidase enhancer 1, which binds to the C-terminal propeptide of type I procollagen; TLL1, Tolloid-like protein 1, protease that processes procollagen C-propeptides; BMP1, bone morphogenetic protein 1, which cleaves the C-terminal propeptides of procollagen I, II, and III. (b) LOXL2 spatial expression in healthy liver cells.
Altered expression and activation of LOX family members can affect the tissue microenvironment, which is implicated in many pathological conditions, including tissue fibrosis, atherosclerosis, and tumor development [53]. Previous studies have indicated that LOXL2 expression is regulated at the transcriptional and post-transcriptional levels [54,55,56], as well as by alternative splicing [57,58], and through an interaction with micro-RNA molecules [59,60,61]. LOXL2 protein activity can also be regulated by post-translational modifications, specifically glycosylation, while an aberrant level of glycosylation has been linked to human malignancies [62]. N-glycosylation is typical in membrane and secretory proteins, while the recombinant human LOXL2 protein, from human embryonic kidney cells, is glycosylated at N288, N455, and N644 (N-linked glycans), which significantly improves its stability and secretion (Figure 1) [63].
In recent years, an increasing number of studies have revealed the upregulation of LOXL2 expression in various solid tumors [64]. However, the mechanism of LOXL2-mediated tumor progression has mostly been investigated in breast cancer [64,65]. Interestingly, in breast cancer cell lines overexpressing LOXL2, two forms of LOXL2 were generated: a non-glycosylated intracellular form (~75-kDa) and an extracellular form (~100-kDa), which had been glycosylated at N455 and N644 [66]. Moreover, it has been shown that breast carcinomas have a more prominent invasive ability, via the actions of intracellular LOXL2, compared to the extracellular effects of LOXL2 in ECM remodeling, thereby suggesting that LOXL2 has specific intra- and extracellular functions [64,65,66]. Tumor progression induced by enhanced LOXL2 expression has been well documented in various digestive system-related cancers, including HCC [23]; thus, its extra- and intracellular functions should be further explored as a potential novel therapeutic target for HCC.
3. LOXL2 Expression in HCC and Correlation with Clinical Parameters
LOXL2 is overexpressed in human HCC tissue compared to healthy liver tissue, at both the mRNA and protein levels [17,24,67,68,69,70,71,72]. LOXL2 protein expression has been shown to correlate to the amount of fibrosis in the tumor stroma and was more pronounced in the cytoplasm of cancer cells directly adjacent to a fibrous stroma, compared to centrally located cancer cells [71]. LOXL2 expression was positively correlated with the direct invasion of adjacent liver tissue [17], increased frequent portal vein invasion, poor tumor differentiation, and more advanced TNM stage [71]. Moreover, LOXL2 expression in HCC patients, especially high-risk HCCs (HCC tumor size > 5 cm, HCCs with portal vein invasion, poor differentiation, and TNM stage II or III), correlated with shorter overall survival, disease-free survival, disease-specific survival, and extrahepatic recurrence-free survival [24,69,70,71]. Hypoxia, chronic inflammation, and fibrosis have all been shown to induce LOXL2 expression in HCC [15,17]. Additionally, in human HCC tissue samples, the co-expression of LOXL2 and carbon anhydrase IX, which is a hypoxia-related biomarker, showed a better correlation with prognostic parameters compared to LOXL2 alone [71]. Interestingly, LOXL2 levels were significantly higher in the serum of HCC patients than in the serum of non-HCC patients [17], making LOXL2 a promising biomarker for HCC.
4. LOXL2 and micro-RNAs in HCC
Recent studies have revealed that micro-RNAs (miRs) may be differentially expressed in HCC and that some of them function as negative epigenetic regulators of LOXL2, thereby affecting the LOXL2 TME-mediated processes. Wong et al. demonstrated that miR-26 and miR-29 suppress LOXL2 in HCC by directly binding to the 3′UTR of LOXL2 mRNA [17]. Specifically, miR29-a has been identified as a negative regulator of hypoxia-responsive genes, such as HIF-1α, VEGFa, LOX, and LOXL2 in HCC cell lines and HCC tissue functional studies [69,136]. Repression of miR-29a has been mediated by MYC; MYC amplification represents one of the earliest events in HCC development [137,138]. Recent bioinformatics analysis proposed hsa-miR-192-5p as a potential LOXL2 regulator in HCC; however, more in vivo studies are required to clarify these interactions between LOXL2 and specific micro-RNAs in the pathogenesis of HCC [139].
5. LOXL2 as Potential Target for Treatment of HCC
The well-established treatment options for patients with HCC have limited clinical efficacy in the late stages of HCC due to severe fibrosis and cirrhosis [140]. Therefore, chemoresistant HCC cases require novel combined treatment strategies. Since LOXL2 is unquestionably involved in almost every step of HCC progression and dissemination, this protein fulfills conditions that are required for a potential target in developing molecular anticancer therapy. Considering the LOXL2 protein structure, two approaches for targeting this protein were considered in drug development studies. The first was based on the fact that LOXL2 demands copper ions for enzyme activity, and the second was to target the lysine tyrosylquinone region (LTQ), which possesses a cofactor binding function [141]. Designing highly selective LOXL2 inhibitors is promising and could be beneficial for HCC patients with LOXL2 overexpression since the downregulation of LOXL2 expression reduces tumor cell invasiveness and metastatic spread [142].
The first monoclonal LOXL2 antibody, AB0023, which specifically binds to the fourth SRCR domain was developed in 2010 and has been effective in clinical studies (Table 1) [49,127,143]. Its efficacy was demonstrated in both tumor xenografts as well as in liver fibrosis models [49,127]. On the other hand, simtuzumab, a humanized LOXL2 antibody (AB0024), has not achieved satisfactory results in clinical trials for fibrotic diseases and several solid tumors [144,145,146]. The lack of clinical effectiveness can be explained by the fact that specific antibody inhibitors probably only deactivate extracellular LOXL2, while the intracellular functions of LOXL2 remain preserved due to incomplete antibody internalization or the compensatory activity of other LOX family members [25,82]. In the search for more selective LOXL2 inhibitors, most efforts have been toward designing a small molecule inhibitor in order to increase specificity and efficacy, and to minimize its side effects [141]. According to the data available on the PHAROS web interface for exploring target/ligand interactions [147], a query for ”LOXL2” resulted in 227 active ligands (ChEMBL compounds with an activity cutoff of <30 nM), although clinical trials that focus on testing LOXL2 based-drugs for HCC are still lacking.
One of the most explored LOX inhibitors is β-aminopropionitrile (BAPN), a small irreversible inhibitor that blocks the catalytic activity of all LOX members and displays specific affinity to LOXL2 [141,148]. Previous studies have shown significant tumor suppressive effects by BAPN in several solid tumors and tumor-cell lines [149,150,151,152,153]. Moreover, some studies demonstrated that BAPN affects the TME and impedes the interaction between cancer-associated fibroblasts and gastric cancer cells, resulting in a reduction in the frequency and size of the liver metastasis [154]. In HCC, BAPN inhibited tumor growth and angiogenesis in vivo and hampered the migration and invasion of HCC cell lines [70,118]. According to the study by Liu et al. [155], BAPN also showed an ameliorative effect in the CCl4-induced model of liver fibrosis. The first selective LOXL2 inhibitor, LOXL2-IN-1 hydrochloride, demonstrated potential for use in HCC treatment since it suppressed Snail1, HIF-1α, and VEGF, the main drivers of HCC progression, as mentioned above [24]. In recent years, the second generation of small-molecular-weight haloallylamine-based LOXL2 inhibitors was explored, including PXS-5338, PXS-5382, and PXS-5878, which showed promising results in inhibiting the catalytic activity of LOXL2 [156]. A dual LOXL2/LOXL3 inhibitor, PXS-5153A, was designed in 2019 by Schilter et al. [157] and also demonstrated ameliorating effects and a significant improvement in liver fibrosis.
Table 1. LOXL2 inhibitors for HCC and liver fibrosis.
| Type | Agent | Target | References |
|---|---|---|---|
| monoclonal antibody |
AB0023 | LOXL2 | [46,121,122] |
| AB0024 | LOXL2 | [123,124,125] | |
| small-molecule inhibitor |
BAPN | LOX/LOXL1-4 | [100,133,134] |
| LOXL2-IN-1 | LOXL2 | [122] | |
| PXS-5338 | LOXL2 | [156] | |
| PXS-5382 | LOXL2 | [156] | |
| PXS-5878 | LOXL2 | [156] | |
| PXS-5153A | LOXL2/LOXL3 | [157] | |
| (2-chloropyridin-4-yl) methenamine | LOXL2 | [158] |
Currently, the most promising results on the potential utility of LOXL2 inhibitors in HCC treatment have been published by Gong et al. [15]. The authors demonstrated that a highly selective LOXL2 inhibitor, namely (2-chloropyridin-4-yl) methenamine [158], in combination with 5FU and sorafenib treatment, significantly decreased the tumor size in a mouse model study. Moreover, HCC cells treated with this agent showed a significantly better response to 5FU and sorafenib treatment [15]. These results suggest that LOXL2 inhibition has the potential to be more successful in patients with HCC in late fibrotic stages with the concomitant application of 5FU and sorafenib. However, these observations in animal studies need to be validated in further clinical studies.
This entry is adapted from the peer-reviewed paper 10.3390/ijms241411745
This entry is offline, you can click here to edit this entry!