Eriophyoidea are a highly diverse lineage of plant-feeding arthropods that have proven to be elusive with respect to their placement within Acariformes [
1]. Throughout the history of acarology, this group has been shifted around to very different taxonomic positions, including both within and outside Trombidiformes. From the mid-twentieth century until very recently, it was widely thought that Eriophyoidea belong within Trombidiformes, but in the past few years this hypothesis has been greatly undermined by a series of molecular phylogenetic analyses [
2,
3,
4,
5,
6,
7,
8,
9,
10] and a new morphology-based analysis [
11].
2. The Morphological Era (1877–2015)
Two very early classification schemes for mites, which both date to 1877 [
15,
16], placed Eriophyoidea within Acaridae (=Astigmata). This was due, at least in part, to the absence of stigmata. Shortly afterward, in 1884, Claus and Moquin-Tandon [
17] treated Eriophyoidea as a separate and distinct group from all the other main branches of mites recognized at the time. Eriophyoids have also been grouped with Demodicidae in Vermiformia based on their shared worm-like body form [
18], but this placement was not followed in the classification schemes of Oudemans [
19] and Reuter [
20]. Due to a shared feeding ecology, Oudemans suspected that Eriophyoidea were closely affiliated with phytophagous taxa within Trombidiformes. However, Reuter rejected this idea based on the morphology of the digestive system. Trombidiformes and Oribatida have a diverticulum (caecum) comprising a pair of cavities that branch away from the main passage of the gut [
21].
Baker and Wharton [
22] treated Eriophyoidea as one of three basal lineages within Trombidiformes, the others being Tarsonemini and Prostigmata. Somewhat confusingly, they also inferred that eriophyoids share a recent common ancestry with Tetranychidae and Phytoptipalpidae (=Tenuipalpidae), which are in a relatively derived position within Trombidiformes [
4,
5]. They based this inference on the following characters: stylet-like chelicerae, phytophagy, rayed empodia (Eriophyoidea and Tenuipalpidae), and elongate bodies (Eriophyoidea and some Tenuipalpidae) [
22,
23].
It is remarkable that a close relationship between Eriophyoidea and Nematalycidae was never proposed as likely until very recently [
11]. These taxa share a number of similarities that are readily apparent without the need for a detailed morphological analysis, including an annulated and worm-shaped body, unpaired
vi (the rostral seta on the prodorsum) when present, and the absence of prodorsal trichobothria and body lyrifissures (cupules). Keifer [
29] noted that both taxa have genitalia that are relatively anteriorly positioned on a worm-like body. However, he used these similarities, probably assumed by him to be convergent, only to explain the loss of legs III and IV in Eriophyoidea rather than infer a close phylogenetic relationship.
In the period leading up to the relocation of Eriophyoidea to a more basal position within Trombidiformes, the position of Nematalycidae was also in flux. Whereas Nematalycidae were originally thought to belong in Endeostigmata [
31], Cunliffe [
32] hypothesized that this family is more closely related to Tydeoidea, but he provided no arguments to support this hypothesis. In accordance with Cunliffe, in the following decades, most authors placed this family within Tydeoidea [
33,
34] or as a separate superfamily that is allied to Tydeoidea [
35]. Nematalycidae and Tydeoidea share a number of characters that are either plesiomorphic (indirect sperm transfer and the retention of fundamental setae on coxisterna II) or homoplastic (undivided femora, fusion of the palpal femur with the palpal genu). Other characters strongly suggest that Nematalycidae fall outside Trombidiformes. For example, nematalycids clearly bear rutella [
36,
37,
38], structures that are widely thought to be lost throughout Trombidiformes [
36,
39]. Eriophyoids bear infracapitular guides, which are possible homologues of rutella [
38,
40].
Although Kethley treated Nematalycidae as allied to Tydeoidea [
35], soon afterwards he hypothesized that Nematalycidae group with Micropsammidae and Proteonematalycidae within Nematalycoidea [
41]. However, in his discussion of the support for this relationship, the only character he listed that is shared by all three families is the absence of trichobothria. He made no mention of where Eriophyoidea belonged, and there is no evidence that he considered the possibility that Eriophyoidea is affiliated with Nematalycidae.
Lindquist clearly did consider the possibility of a close relationship between Eriophyoidea and Nematalycidae, but he largely rejected this idea in favor of a sister relationship between Eriophyoidea and Tydeoidea [
27]. One of his main reasons for doing so was that Nematalycoidea, which is almost certain to be an artificial group [
4,
11], was hypothesized to be in a basal position, outside of Trombidiformes (largely because of the shared possession of rutella). By this time, Lindquist had also undertaken an unpublished cladistic analysis of Trombidiformes, in which Eriophyoidea was recovered as sister to Tydeoidea (Lindquist, pers. comm. 10 April 2023). A single cladogram from this analysis was published without any associated data [
28].
Therefore, although Nematalycidae were relocated outside Trombidiformes, the same treatment was apparently not considered for Eriophyoidea. This was somewhat remedied by Lindquist when he suggested that Eriophyoidea may be closely affiliated with Alycidae [
40]. However, later classification schemes followed Lindquist’s earlier work [
39,
42].
Lindquist contended that the consensus of morphological evidence favored a sister relationship between Eriophyoidea and Tydeoidea [
27], although many of the characters that he used in support of this relationship show an equal or greater resemblance between Eriophyoidea and Nematalycidae [
11].
Of the characters used by Lindquist [
27] to argue for a sister relationship between Eriophyoidea and Tydeoidea, the only ones that show a greater degree of resemblance between eriophyoids and tydeoids than between eriophyoids and nematalycids are a suite of strongly interdependent and homoplastic characters pertaining to paedomorphisms, including the partial or complete suppression of anamorphosis, the loss of urstigmata, and the suppression of genital papillae and the nymphal progenital chamber [
11].
3. Molecular Era (2016–Present)
The first molecular phylogenetic analysis to address the position of Eriophyoidea was based on a mitogenomic analysis and showed strong support for the placement of Eriophyoidea outside of Trombidiformes [
2]. Subsequent mitogenomic analyses also recovered that result [
6,
7,
8,
9]. Only a single mitogenomic analysis, which is undermined by the undersampling of non-trombidiform taxa, has recovered Eriophyoidea from within Trombidiformes [
13]. Whole-genomic analyses have also recovered Eriophyoidea from outside of Trombidiformes [
3,
10]. Therefore, there is near unanimity among whole-genomic and mitogenomic analyses.
A substantial proportion of phylogenetic analyses provided support for a close relationship between Eriophyoidea and Nematalycidae; Eriophyoidea are either sister to Nematalycidae [
4,
5] or nested within Nematalycidae [
4,
11]. These analyses are based on morphology [
11], Sanger sequencing [
4,
5], and whole genomes [
3]. The whole genome analysis, which is the only phylogenomic analysis to date to have included endeostigmatic taxa, revealed that Eriophyoidea and Nematalycidae share a suite of highly conserved nuclear proteins [
3]. With respect to all the phylogenetic analyses that recovered Eriophyoidea outside of Trombidiformes, their congruence with a close relationship between Eriophyoidea and Nematalycidae is strong support for this relationship and also for the placement of Eriophyoidea outside of Trombidiformes (
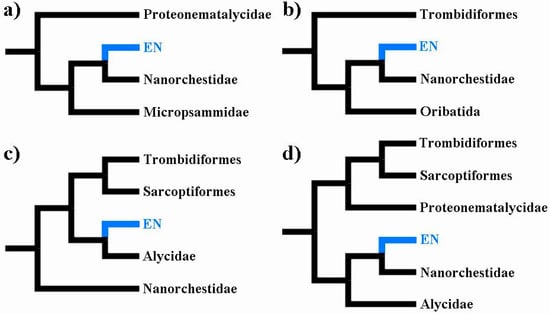
Figure 2).
Figure 2. Summary trees pertaining to analyses that have recovered an Eriophyoidea-Nematalycidae (EN) clade (highlighted blue): (
a) parsimony tree (showing only part of Sarcoptiformes) based on 110 morphological characters [
11]; (
b) maximum likelihood tree (low taxonomic resolution) based on 90 orthologous proteins [
3]; (
c) maximum likelihood tree based on COI, 18S rRNA, 28S rRNA, EF1-α, SRP54, and HSP70; [
5]; (
d) Bayesian time tree based on COI, 18S rRNA, 28S rRNA, and HSP70 [
4]. Across all trees, Nematalycidae were recovered as paraphyletic (
a) or sister (
c,
d) to Eriophyoidea—taxonomic sampling was insufficient to resolve this for the 90 orthologous proteins (whole genome data) (
b).
On the other hand, there is very little consistency in the results of analyses that have recovered Eriophyoidea from within Trombidiformes. Eriophyoidea have been found in the following positions: (1) sister to a clade comprising Tydeoidea, Eupodidae, and Adamystidae [5: 18S rRNA, 28S rRNA, EF1-α, SRP54, HSP70, and COI]; (2) sister to a clade comprising Parasitengona, Cheyletoidea, and Eupodides [13: mitochondrial genomes]; (3) nested within Tydeoidea [12: 18S rRNA, 28S rRNA, and COI]; (4) sister to Tydeoidea [14: 12S rRNA, 18S rRNA, 28S rRNA, and COI]; (5) sister to a clade comprising Nematalycidae and Benoinyssus (a genus within Eupodidae) [14: 12S rRNA, 18S rRNA, 28S rRNA, and COI]. The incongruences among these results undermine the case for the placement of Eriophyoidea within Trombidiformes.
4. Are Eriophyoidea Nested within Tydeoidea?
More detailed consideration will now be given to the results of Szudarek-Trepto et al. [
12] (from here on abbreviated to ST22) because they recently recovered Eriophyoidea as nested within Tydeoidea, which is a similar result to the sister relationship that was hypothesized for the two taxa [
26,
27,
28]. The molecular phylogeny of ST22 was based on only three loci (18S rRNA, 28S rRNA, and COI), and the results are undermined by a failure to account for the rRNA secondary structure (rDNA stem complementarity) as the basis of alignment. Moreover, ST22 undertook a manual alignment for genera and families without testing for their monophyly first, which represents a form of circular reasoning (an alignment favoring traditional groupings can skew an analysis toward a traditional phylogeny).
Hypervariable regions experience frequent expansion–contraction and instability in corresponding stem-loop rRNA structures, so sequence homology cannot be established here, except for closely related taxa [
47,
48,
49]. This major methodological flaw further undermines the results of ST22. When attempting to undertake a phylogenetic analysis to resolve internal relationships within Phytoptidae (Eriophyoidea), Chetverikov et al. [
50] had to exclude the hypervariable regions because they were unalignable.
ST22 also recovered Eriophyoidea nested within Tydeoidea based on a separate morphology-based analysis. Unfortunately, they omitted Nematalycidae (and any other Endeostigmata), thereby disregarding alternative hypotheses. Only four superfamilies were included in the ingroup, thus greatly constraining the morphological analysis to produce the same result as the molecular analysis. ST22 claim that eight characters “unambiguously define” an Eriophyoidea-Tydeoidea clade. However, only two of these characters are synapomorphies (characters #1, #2), and the remaining six are homoplasies.
There are also other apparent weaknesses in the coding. The third character (#3) addresses the potential absence of cheliceral setae
chb in Eriophyoidea and Tydeoidea. However, the absence of
chb in Eriophyoidea cannot be asserted with confidence ([
11,
51] (Figure 5D,E) and [
52]). Secondly, in both Eriophyoidea and Tydeoidea, setae
ps3 (the third pair of setae on the pseudanal (
PS) segment) were coded as absent (character #41), but in Eriophyoidea the absence of
ps3 is attributable to the hypothesized absence of the pseudanal segment, whereas the absence of
ps3 in Tydeidae is not. Setae
ps3 should have therefore been coded as unknown or “not applicable” in Eriophyoidea. The miscoding of this character generated the only morphological support for the nested position of Eriophyoidea within Tydeoidea.
With the single exception of the absence of urstigmata, the homoplasious character states used by ST22 to support an Eriophyoidea-Tydeoidea clade (absence of naso; absence of urstigmata; undivided femur I; undivided femur II; undivided femur III, but note that this is inapplicable to Eriophyoidea; absence of rhagidial organs) are also shared with Nematalycidae [
11].
Of all the different and competing hypotheses on the position of Eriophyoidea, a nested placement within Tydeoidea must be one of the least likely because it is incongruent with the fundamentally different modes of gnathosomal integration of these two taxa. Gnathosomal integration proceeded in Tydeoidea via the reduction of the fixed digits, so that a cheliceral groove formed along the dorsum of the subcapitulum around the movable digits [
53]. This reduction circumvented the need for the modification of any part of the subcapitulum into a sheath, as instead occurred in Eriophyoidea and some Nematalycidae [
38].