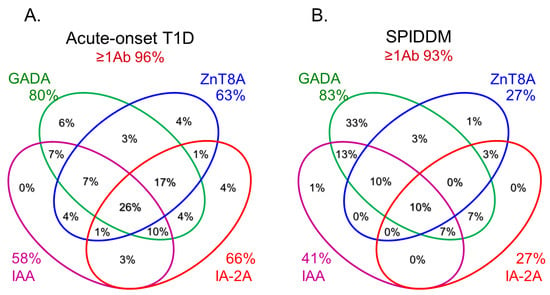
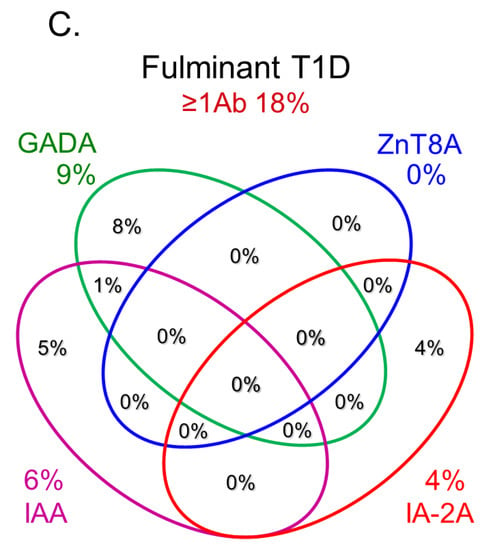
Anti-islet autoantibodies serve as key markers in immune-mediated type 1 diabetes (T1D) and slowly progressive T1D (SPIDDM), also known as latent autoimmune diabetes in adults (LADA). Autoantibodies to insulin (IAA), glutamic acid decarboxylase (GADA), tyrosine phosphatase-like protein IA-2 (IA-2A), and zinc transporter 8 (ZnT8A) are currently employed in the diagnosis, pathological analysis, and prediction of T1D. GADA can also be detected in non-diabetic patients with autoimmune diseases other than T1D and may not necessarily reflect insulitis. Conversely, IA-2A and ZnT8A serve as surrogate markers of pancreatic β-cell destruction. A combinatorial analysis of these four anti-islet autoantibodies demonstrated that 93–96% of acute-onset T1D and SPIDDM cases were diagnosed as immune-mediated T1D, while the majority of fulminant T1D cases were autoantibody-negative.
- enzyme-linked immunosorbent assay
- epitope
- glutamic acid decarboxylase
- Latent-Autoimmune Diabetes in Adults (LADA)
- Slowly-progressive type 1 diabetes (SPIDDM)
1. Introduction
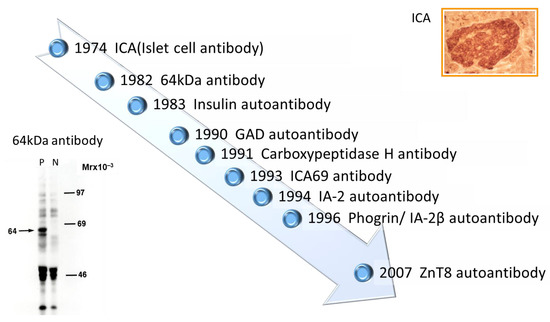
2. History of Anti-Islet Autoantibody Discovery
| Name of Antigen | Localization | Function | Reference |
|---|---|---|---|
| Insulin | Insulin secretory granules | Regulate glucose levels in the blood and induce glucose storage in the liver, muscles, and adipose tissue | [7] |
| GAD65 | Synaptic-like vesicles in the cytoplasm of β-cells | Rate-limiting enzyme engaged in the synthesis of the neurotransmitter γ-aminobutyric acid from L-glutamate | [8] |
| GAD67 | Cytosol of β-cells | Rate-limiting enzyme engaged in the synthesis of the neurotransmitter γ-aminobutyric acid from L-glutamate | [11] |
| IA-2 | Insulin secretory granule membrane | Regulate insulin secretory granule content and β-cell growth | [9,12] |
| Phogrin/IA-2β | Insulin secretory granule membrane | Regulate insulin secretory granule content and β-cell growth | [13,14] |
| Carboxypeptidase H | Insulin secretory granules and granule membrane | Convert proinsulin into insulin and C-peptide by catalyzing the release of C-terminal arginine or lysine residues from polypeptides | [15] |
| ICA69 | Insulin secretory granule membrane | Dense-core vesicles signaling and maturation | [16] |
| ZnT8 | Insulin secretory granule membrane | Transport zinc ion from the cytosol into the insulin secretory granules | [17,18] |
| GM2-1 ganglioside | Secretory granules in β-cells and non-β-cells | unknown | [19] |
| Heat shock protein 60 | Insulin secretory granules | Assist correct folding of partially folded polypeptides and presentation of antigen to MHC molecules | [20] |
| GLUT2 | β-cell surface membrane | Uptake glucose from the blood into β-cells | [21] |
| Tetraspanin-7 | Insulin secretory granule membrane | Regulate Ca2+-dependent insulin exocytosis | [22] |
| ICA12/SOX13 | Cytoplasm and nucleus in β-cells and non-β-cells | Transcription factor (Function in the islets is unknown) | [23] |
3. Pathophysiology of the Generation of Anti-Islet Autoantibodies
4. Localization and Function of Autoantigens against Anti-Islet Autoantibodies
- (1)
-
Insulin
- (2)
-
GAD 65 and GAD67
- (3)
-
IA-2 and IA-2β/phogrin
- (4)
-
ZnT8
- (5)
-
Carboxypeptidase H
- (6)
-
ICA69
- (7)
-
GM2-1 ganglioside
- (8)
-
Heat shock protein 60
- (9)
-
GLUT2
- (10)
-
Tetraspanin-7
- (11)
-
ICA12/SOX13
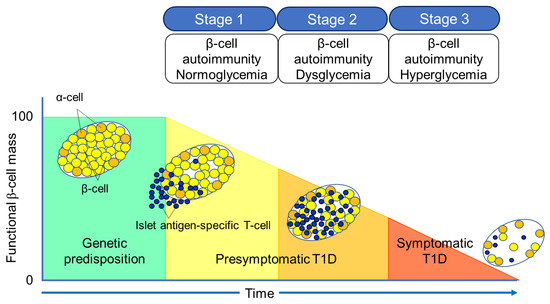
5. Significance of Anti-Islet Autoantibodies in the Pathophysiology of T1D
| Subject | Prevalence |
|---|---|
| Healthy control | <1% |
| Acute-onset type 1 diabetes (at onset) | 60–80% |
| Fulminant type 1 diabetes | 5–9% |
| LADA (SPIDDM) | 100% |
| Type 2 diabetes (diet/OHA) | 4–5% |
| Polyglandular autoimmune syndrome, type 1 | 30–40% |
| Polyglandular autoimmune syndrome, type 2 | 30–50% |
| Autoimmune thyroid disease | 6–8% |
| Stiff-person syndrome | 60–70% |
6. Role of Anti-Islet Autoantibodies in the Diagnosis of T1D
References
- ElSayed NA, Aleppo G, Aroda VR, Bannuru RR, Brown FM, Bruemmer D, Collins BS, Hilliard ME, Isaacs D, Johnson EL, Kahan S, Khunti K, Leon J, Lyons SK, Perry ML, Prahalad P, Pratley RE, Seley JJ, Stanton RC, Gabbay RA, on behalf of the American Diabetes Association. Classification and diagnosis of diabetes: Standards of care in diabetes-2023. Diabetes Care 46(Supplement_1): S19-S40, 2023
- Roep BO, Thomaidou S, van Tienhoven R, Zaldumbide A. Type 1 diabetes mellitus as a disease of the β-cell (do not blame the immune system?). Nat Rev Endocrinol. 17:150-161, 2021
- Karvonen M, Viik-Kajander M, Moltchanova E, Libman I, LaPorte R, Tuomilehto J. Incidence of childhood type 1 diabetes worldwide. Diabetes Mondiale (DiaMond) Project Group. Diabetes Care. 23:1516-26, 2000
- Kawasaki E, Matsuura N, Eguchi K. Type 1 diabetes in Japan. Diabetologia. 49:828-836, 2006
- Bottazzo GF, Florin-Christensen A, Doniach D. Islet-cell antibodies in diabetes mellitus with autoimmune polyendocrine deficiencies. Lancet. 2(7892):1279-1283, 1974
- Baekkeskov S, Nielsen JH, Marner B, Bilde T, Ludvigsson J, Lernmark Å. Autoantibodies in newly diagnosed diabetic children immunoprecipitate human pancreatic islet cell proteins. Nature. 298(5870):167-169, 1982
- Palmer JP, Asplin CM, Clemons P, Lyen K, Tatpati O, Raghu PK, Paquette TL. Insulin antibodies in insulin-dependent diabetics before insulin treatment. Science. 222(4630):1337-9, 1983
- Baekkeskov S, Aanstoot HJ, Christgau S, Reetz A, Solimena M, Cascalho M, Folli F, Richter-Olesen H, De Camilli P. Identification of the 64K autoantigen in insulin-dependent diabetes as the GABA-synthesizing enzyme glutamic acid decarboxylase. Nature. 347(6289):151-156, 1990
- Christgau S, Schierbeck H, Aanstoot HJ, Aagaard L, Begley K, Kofod H, Hejnaes K, Baekkeskov S. Pancreatic beta cells express two autoantigenic forms of glutamic acid decarboxylase, a 65-kDa hydrophilic form and a 64-kDa amphiphilic form which can be both membrane-bound and soluble. J Biol Chem. 266:21257-21264, 1991
- Lan MS, Lu J, Goto Y, Notkins AL. Molecular cloning and identification of a receptor-type protein tyrosine phosphatase, IA-2, from human insulinoma. DNA Cell Biol. 13:505-14, 1994
- Harashima S, Clark A, Christie MR, Notkins AL. The dense core transmembrane vesicle protein IA-2 is a regulator of vesicle number and insulin secretion. Proc Natl Acad Sci USA. 102:8704-8709, 2005
- Kawasaki E, Hutton JC, Eisenbarth GS. Molecular cloning and characterization of the human transmembrane protein tyrosine phosphatase homologue, phogrin, an autoantigen of type 1 diabetes. Biochem Biophys Res Commun. 227:440-447, 1996
- Cai T, Hirai H, Zhang G, Zhang M, Takahashi N, Kasai H, Satin LS, Leapman RD, Notkins AL. Deletion of IA-2 and/or IA-2β in mice decreases insulin secretion by reducing the number of dense core vesicles. Diabetologia. 54:2347-2357, 2011
- Aguilar-Diosdado M, Parkinson D, Corbett JA, Kwon G, Marshall CA, Gingerich RL, Santiago JV, McDaniel ML. Potential autoantigens in IDDM. Expression of carboxypeptidase-H and insulin but not glutamate decarboxylase on the β-cell surface. Diabetes. 43:418-425, 1994
- Spitzenberger F, Pietropaolo S, Verkade P, Habermann B, Lacas-Gervais S, Mziaut H, Pietropaolo M, Solimena M. Islet cell autoantigen of 69 kDa is an arfaptin-related protein associated with the Golgi complex of insulinoma INS-1 cells. J Biol Chem. 278:26166-26173, 2003
- Dwivedi OP, Lehtovirta M, Hastoy B, Chandra V, Krentz NAJ, Kleiner S, Jain D, Richard AM, Abaitua F, Beer NL, Grotz A, Prasad RB, Hansson O, Ahlqvist E, Krus U, Artner I, Suoranta A, Gomez D, Baras A, Champon B, Payne AJ, Moralli D, Thomsen SK, Kramer P, Spiliotis I, Ramracheya R, Chabosseau P, Theodoulou A, Cheung R, van de Bunt M, Flannick J, Trombetta M, Bonora E, Wolheim CB, Sarelin L, Bonadonna RC, Rorsman P, Davies B, Brosnan J, McCarthy MI, Otonkoski T, Lagerstedt JO, Rutter GA, Gromada J, Gloyn AL, Tuomi T, Groop L. Loss of ZnT8 function protects against diabetes by enhanced insulin secretion. Nat Genet 51, 1596-1606, 2019
- Nicolson TJ, Bellomo EA, Wijesekara N, Loder MK, Baldwin JM, Gyulkhandanyan AV, Koshkin V, Tarasov AI, Carzaniga R, Kronenberger K, Taneja TK, da Silva Xavier G, Libert S, Froguel P, Scharfmann R, Stetsyuk V, Ravassard P, Parker H, Gribble FM, Reimann F, Sladek R, Hughes SJ, Johnson PR, Masseboeuf M, Burcelin R, Baldwin SA, Liu M, Lara-Lemus R, Arvan P, Schuit FC, Wheeler MB, Chimienti F, Rutter GA. Insulin storage and glucose homeostasis in mice null for the granule zinc transporter ZnT8 and studies of the type 2 diabetes-associated variants. Diabetes 58, 2070-2083, 2009
- Dotta F, Previti M, Neerman-Arbez M, Dionisi S, Cucinotta D, Lenti L, Di Mario U, Halban PA. The GM2-1 ganglioside islet autoantigen in insulin-dependent diabetes mellitus is expressed in secretory granules and is not β-cell specific. Endocrinology. 139:316-319, 1998
- Ozawa Y, Kasuga A, Nomaguchi H, Maruyama T, Kasatani T, Shimada A, Takei I, Miyazaki J, Saruta T. Detection of autoantibodies to the pancreatic islet heat shock protein 60 in insulin-dependent diabetes mellitus. J Autoimmun. 9:517-524, 1996
- Inman LR, McAllister CT, Chen L, Hughes S, Newgard CB, Kettman JR, Unger RH, Johnson JH. Autoantibodies to the GLUT-2 glucose transporter of β cells in insulin-dependent diabetes mellitus of recent onset. Proc Natl Acad Sci USA. 90:1281-1284, 1993
- Walther D, Eugster A, Jergens S, Gavrisan A, Weinzierl C, Telieps T, Winkler C, Ziegler AG, Bonifacio E. Tetraspanin 7 autoantibodies in type 1 diabetes. Diabetologia. 59:1973-1976, 2016
- Kasimiotis H, Myers MA, Argentaro A, Mertin S, Fida S, Ferraro T, Olsson J, Rowley MJ, Harley VR. Sex-determining region Y-related protein SOX13 is a diabetes autoantigen expressed in pancreatic islets. Diabetes. 49:555-561, 2000
- Wenzlau JM, Juhl K, Yu L, Moua O, Sarkar SA, Gottlieb P, Rewers M, Eisenbarth GS, Jensen J, Davidson HW, Hutton JC. The cation efflux transporter ZnT8 (Slc30A8) is a major autoantigen in human type 1 diabetes. Proc Natl Acad Sci USA. 104:17040-17045, 2007
- Fousteri G, Ippolitoa E, Ahmedb R, Hamad ARA. Beta-cell specific autoantibodies: Are they just an indicator of type 1 diabetes? Curr Diabetes Rev. 13: 322-329, 2017
- Mallone R, Brezar V. To B or not to B: (anti)bodies of evidence on the crime scene of type 1 diabetes? Diabetes 60:2020-2022, 2011
- Pinto AI, Smith J, Kissack MR, Hogg KG, Green EA. Thymic B cell-mediated attack of thymic stroma precedes type 1 diabetes development. Front Immunol 9:1281, 2018
- Insel RA, Dunne JL, Atkinson MA, Chiang JL, Dabelea D, Gottlieb PA, Greenbaum CJ, Herold KC, Krischer JP, Lernmark Å, Ratner RE, Rewers MJ, Schatz DA, Skyler JS, Sosenko JM, Ziegler AG. Staging presymptomatic type 1 diabetes: a scientific statement of JDRF, the Endocrine Society, and the American Diabetes Association. Diabetes Care. 38:1964-1974, 2015
- Ziegler AG, Rewers M, Simell O, Simell T, Lempainen J, Steck A, Winkler C, Ilonen J, Veijola R, Knip M, Bonifacio E, Eisenbarth GS. Seroconversion to multiple islet autoantibodies and risk of progression to diabetes in children. JAMA 309: 2473-2479, 2013
- Krischer JP; Type 1 Diabetes TrialNet Study Group. The use of intermediate endpoints in the design of type 1 diabetes prevention trials. Diabetologia 56: 1919-1924, 2013
- Verge CF, Gianani R, Kawasaki E, Yu L, Pietropaolo M, Jackson RA, Chase HP, Eisenbarth GS.Prediction of type 1 diabetes in first-degree relatives using a combination of insulin, GAD, and ICA512bdc/IA-2 autoantibodies. Diabetes. 45:926-933, 1996
- Yu L, Rewers M, Gianani R, Kawasaki E, Zhang Y, Verge C, Chase P, Klingensmith G, Erlich H, Norris J, Eisenbarth GS. Antiislet autoantibodies usually develop sequentially rather than simultaneously. J Clin Endocrinol Metab. 81(12):4264-4267, 1996
- Nakamura K, Kawasaki E, Abiru N, Jo O, Fukushima K, Satoh T, Kuriya G, Kobayashi M, Kuwahara H, Yamasaki H, Ide T, Eguchi K. Trajectories of anti-islet autoantibodies before development of type 1 diabetes in interferon-treated hepatitis C patients. Case reports and a literature review. Endocr J. 57:947-951, 2010
- Kawasaki E, Yu L, Rewers MJ, Hutton JC, Eisenbarth GS. Definition of multiple ICA512/phogrin autoantibody epitopes and detection of intramolecular epitope spreading in relatives of patients with type 1 diabetes. Diabetes. 47:733-742, 1998
- Kawasaki E, Yasui J, Tsurumaru M, Takashima H, Ikeoka T, Mori F, Akazawa S, Ueki I, Kobayashi M, Kuwahara H, Abiru N, Yamasaki H, Kawakami A. Sequential elevation of autoantibodies to thyroglobulin and glutamic acid decarboxylase in type 1 diabetes. World J Diabetes. 4:227-30, 2013
- Kawasaki E, Nakamura K, Kuriya G, Satoh T, Kobayashi M, Kuwahara H, Abiru N, Yamasaki H, Matsuura N, Miura J, Uchigata Y, Eguchi K.Differences in the humoral autoreactivity to zinc transporter 8 between childhood- and adult-onset type 1 diabetes in Japanese patients. Clin Immunol. 138:146-153, 2011
This entry is adapted from the peer-reviewed paper 10.3390/ijms241210012