Hybrid breeding is an effective way to increase rice yield, and an
indica intra-subspecies hybrid can increase the yield by 20% compared to conventional rice [
1], accounting for half of the rice planting area and 60% of the total rice production in China [
2]. In the past, hybrid rice mainly utilized hybrid vigour between
indica rice varieties, but it has reached a bottleneck and further yield improvements are difficult to achieve due to the limitations of the genetic diversity in hybrid parent lines [
3]. The
indica and
japonica rice varieties are two main subspecies differentiated during the domestication process of Asian cultivated rice. Hybridization between
indica and
japonica subspecies has stronger heterosis, and the yields of
indica–
japonica hybrids are expected to be 30% higher than
indica intra-subspecies hybrid rice [
1]. However, the utilization of
indica–
japonica heterosis is restricted by many factors, such as non-compatibility of the hybrids, different growth periods, super-parental late maturity of the hybrid offspring, and non-overlapping diurnal flower-opening times (DFOTs), which have led to a stagnation of
indica–
japonica hybrid breeding [
4,
5]. With the cloning of multiple
indica–
japonica hybrid sterility genes,
indica–
japonica hybridization has become possible [
3,
6]. Generally,
japonica rice is used as a sterile line, while
indica rice is used as a restorer line. Usually, the DFOT of
japonica rice is after 12 noon, while that of
indica rice is earlier, usually around 10 a.m. [
7]. The different DFOTs of parental lines significantly reduce the cross-pollination rate and yield, increasing the cost of hybrid seed production, which has become one of the main limiting factors in the development of
indica–
japonica hybrid rice breeding [
7,
8]. Therefore, developing sterile
japonica rice varieties with an early DFOT through genetic methods can increase hybrid seed production and reduce seed production costs.
2. Advances in Research on Rice DFOT
2.1. The Structural and the Physiological Basis of Rice Flower Opening
Rice flower opening involves a series of structural and physiological changes, which have an impact on the DFOT. Therefore, studying the structural and physiological basis of rice flower opening is crucial for understanding the DFOT of rice.
2.1.1. The Structural Basis of Rice Flower Opening
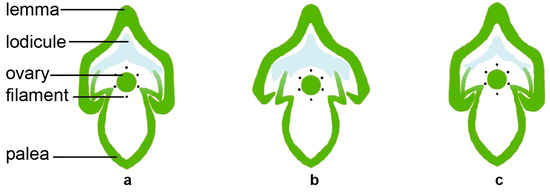
The normal structure of a rice spikelet consists of a pair of sterile lemmas, lemma, palea, a pair of lodicules, six stamens, and a pistil (
Figure 1) [
22]. The lemma and palea of rice enclose the inner floral organs through interlocking grooves. The two lodicules are symmetrically distributed at the base of the ovary adjacent to the lemma; they are composed of large thin-walled cells and evenly distributed small vascular bundles [
4].
Figure 1. The spikelet structure in rice. The morphological anatomy (a) and the cross-section (b) of the spikelet in rice. The red arrows point to the interlocking grooves. Bars: 1 mm.
The lodicule is the most critical organ that controls the opening and closing of the rice spikelet. The opening and closing of the lemma and palea are the processes of the two lodicules absorbing water and swelling or losing water and shrinking (
Figure 2). After the two lodicules absorb water and expand, they push the lemma outward and squeeze the palea inward, causing the interlocking groove of the lemma and palea to loosen, resulting in the opening of the spikelet. When the spikelet is fully opened, the vacuolar membrane of the lodicule cells ruptures, then the cells self-digests its intracellular protein components and organelles. The water and disintegrated substances in the lodicules are transported to the spikelet axis through the vascular bundles, causing the lodicules to lose water. The elasticity of the lemma and palea causes them to close again due to the force of the spikelet axis [
4,
8,
23]. The water absorption and swelling of the lodicules during this process are affected by the degree of relaxation of the lodicule cell wall and the osmotic potential of the cell. Only when the osmotic potential is low and the cell wall is relaxed does the cell have the potential to absorb water and expand. The cell wall plays an important role in supporting and defining the shape of the plant cells and maintaining the cell’s turgor pressure [
24,
25]. In order to achieve the desired size, the plant cells can increase their osmotic pressure and reduce the expansion pressure of the cell wall [
25,
26]. The lodicule is mainly composed of thin-walled cells, which only contain primary cell walls. The primary cell wall is a dynamic structure composed of cellulose, hemicellulose, and pectin, which can be adjusted according to the needs of cell growth and development [
25,
27,
28,
29,
30]. Studies have shown that the degree of methyl-esterification of pectin in the cell wall is positively correlated with the DFOT. Higher pectin methylesterase (PME) activity reduces the degree of methyl-esterification of pectin, increases the degree of calcium binding, makes the cell wall harder, and delays the DFOT. Conversely, lower PME activity increases the degree of methyl-esterification of pectin, reduces the degree of calcium binding, makes the cell wall softer, and advances the DFOT [
31]. Pectin can change the extensibility of the cell wall and may also play a critical role in cell wall remodeling [
30,
32,
33].
Figure 2. The dynamic process of flower opening and lodicule size in the cross-section of rice spikelet. Unopened flower (a), opening flower (b), and opened flower (c).
2.1.2. The Physiological Basis of Flower Opening in Rice
Rice flower opening is the result of a balance between the osmotic pressure and cell wall expansion pressure of the lodicule cells; changes in the metabolites in these cells are crucial for maintaining osmotic pressure. During rice flower opening, the dry weight and soluble sugar content of the lodicule cells increased, resulting in an increase in the osmotic pressure and cell swelling [
4,
34]. Before flower opening, there is an increase in glycolysis activity within the lodicule cells, leading to the appearance of vacuolar invertase, which can hydrolyze sucrose into glucose and fructose, causing an increase in the osmotic pressure within the cells. In addition, during rice flower opening, the expression of the starch hydrolase genes increases, leading to a significant degradation of starch granules in the interlocking groove and lodicules [
35]. This suggests that the increase of soluble sugar content in the lodicules is closely related to glycolysis, vacuolar invertase, and starch hydrolase. The levels of K
+, Na
+, Ca
2+, and Mg
2+ in the lodicules does not change significantly before or during flower opening, and only decreases after the lodicules began to wither. Moreover, the levels of inorganic ions in the lodicules are much lower than those of soluble sugars, suggesting that inorganic ions play a minor role in regulating the osmotic pressure [
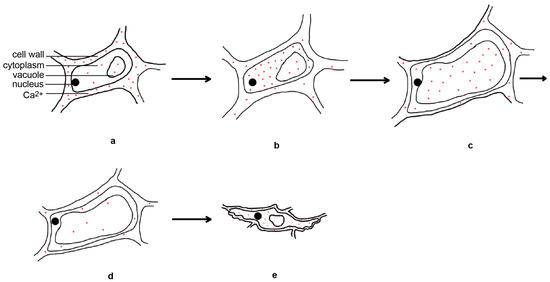
4]. However, another study found that the expansion and withering of the lodicules are related to the spatial and temporal dynamics of Ca
2+ in the lodicules [
36]. The day before flower opening, calcium is mainly located in the cell walls of the lodicule epidermal cells. Four hours before flower opening, the calcium disappears from the cell walls and relocates to the cytoplasm of the lodicule epidermal cells. During flower opening, large amounts of calcium are present in the vacuoles of the lodicule epidermal cells. One hour after flower opening, the calcium content decreases. Six hours after flower opening, calcium accumulates in large quantities in the cytoplasm, vacuole membrane, and cell walls of the lodicule epidermal cells again [
36] (
Figure 3). This indicates that inorganic salt ions may primarily function as signaling molecules in the regulation of flower opening.
Figure 3. Temporal and spatial dynamical changes of calcium ions in lodicule epidermal cells during rice flower opening and closing processes. The day before flower opening (a), four hours before flower opening (b), during flower opening (c), one hour after flower opening (d), and six hours after flower opening (e).
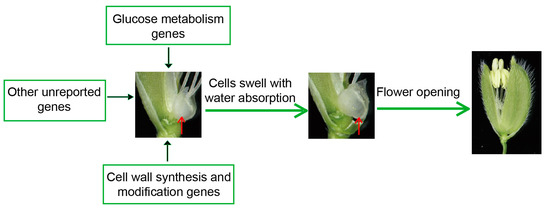
Based on the findings above, we summarized some possible genes affecting the DFOT of rice, including sugar metabolism and cell wall synthesis and modification-related genes (Figure 4). These genes affect the DFOT mainly by changing the expansion pressure or osmotic pressure of the lodicule cells to regulate their size, thereby affecting rice flower opening.
Figure 4. The regulatory mechanism of rice flower opening. The red arrows point to lodicules.
2.2. The Factors Affecting the Regulation of DFOT
2.2.1. The Influence of Genetic Factors on DFOT
Different rice varieties may have different DFOTs [
23]. Generally, the DFOT of the
indica rice is earlier than that of
japonica rice, while the DFOT of wild rice is earlier than that of cultivated rice; this is related to their genetic factors. The DFOT of rice is a stable genetic trait, and the genetic factors that cause differences in DFOT mainly include floral organ characteristics, such as the lodicule structure, grain length and width, number of leaf hairs, and length and softness of glume hairs. Compared with male-fertile rice, male-sterile rice lines usually have a dispersed DFOT, and the peak flower opening period is concentrated in the afternoon. Most flower organs of male-sterile rice lines are deformed, and can include abnormal stamens, degraded male organs, and fewer vascular bundles and conduits in the lodicules. This slows water absorption and reduces the elasticity of the lodicules, leading to a delayed DFOT. As a result, hybrid seed production rates are low due to the parents’ non-overlapping DFOTs during seed production [
37]. There are also differences between the DFOT of
indica and
japonica rice. Among the different grain types in the
indica rice subspecies, the rounder the grain is, the later its DFOT; conversely, the longer and thinner the grain is, the earlier its DFOT [
38]. Long-grain or medium-grain rice varieties, as well as some late
japonica round-grain rice lines show early DFOT characteristics due to their genetic background of
indica rice. Therefore, grain length and width have become one of the reference indicators for selecting male-sterile lines with early DFOT traits [
39]. It is worth noting that there is a certain correlation between rice DFOT and the number of leaf hairs, glume hair characteristics, and seed staining degree with phenol [
40]. The rice varieties with fewer leaf hairs or seeds that are easily stained with phenol will reach the peak flower opening time later, and the varieties with longer, more disordered, and softer glume hairs generally show earlier DFOTs and have a longer diurnal flower opening duration. It is still not clear whether there is a relationship between the DFOT and grain length and leaf hair number. In general,
indica rice has longer grains and more leaf hairs, while
japonica rice has shorter grains and fewer leaf hairs. Generally, the DFOT of
indica rice is much earlier than
japonica rice; therefore, the correlation between the DFOT and the grain length or leaf hair numbers might be due to the differences in the genetic background of
indica and
japonica rice varieties.
2.2.2. The Influence of Plant Hormones and Growth Regulators on DFOT
Plant hormones play an important regulatory role in the opening and closing of rice spikelets. With the development of biotechnology and the increasing demand for hybrid rice seed production, a growing number of plant growth regulators that have been synthesized artificially or are extracted from microorganisms that have similar physiological and biological effects to endogenous plant hormones are being used to solve the problem of non-overlapping DFOTs between the parental lines. Currently, the reported plant growth regulators used to regulate flower opening mainly include methyl jasmonate (MeJA), auxins, “920”, triacontanol, Huaxinling, and others.
Jasmonates (JAs) are a type of fatty acid derivative containing a cyclopentanone basic structure, including jasmonic acid and its various derivatives, such as jasmonoyl-isoleucine (JA-Ile), MeJA, 12-oxo-phytodienoic acid (OPDA), and so on. JAs play an important role in plant growth and development [
41,
42,
43]. The distribution of endogenous JAs in spikelet organs is tissue-specific and development stage-specific. The JA content in rice florets remains stable before flower opening, but sharply increases to a peak during flower opening before declining after flower opening [
44]. Before flower opening, the JA level in the pedicel of the spikelet is the highest. During flower opening, the JA levels in the stamens and lodicules are significantly higher than in other floral organs, while the JA content in the pedicel is the lowest. Consistent with the changes in JA levels, the expression of JA biosynthesis-related genes
OsDAD1,
OsAOS1,
OsAOC,
OsJAR1, and
OsOPR7 in the stamens and lodicules increases significantly, while their expression in the pedicel is significantly down-regulated. The
OsOPR7 gene has been reported to affect carbohydrate transport in rice lodicules during flower opening [
45,
46]. The expression of JA signal transduction pathway-related genes
OsCOI1b and
OsJAZs also increases significantly in the lodicules [
45]. The reduction in JA content in the lodicules of the cytoplasmic male sterile (CMS) line Zhenshan 97A delayed and dispersed the DFOT of spikelets, indicating that endogenous JA regulates the opening and closing process of rice spikelets [
8]. MeJA is an important DFOT regulator in hybrid rice seed production. Spraying MeJA can induce spikelet opening in
japonica CMS lines. The effects of MeJA on inducing flower opening in CMS lines are more sensitive than in fertile lines [
47,
48]. MeJA can also induce flower opening in
indica CMS lines with a significant flower opening peak, showing that MeJA has universal applicability in regulating DFOT in hybrid rice production [
49]. Exogenous MeJA has also been reported to promote the DFOT of sorghum, and the promotion effect increases with the increasing concentration of exogenous MeJA [
50]. Therefore, JAs play an important role in crop breeding.
Auxin has been reported to be associated with plant flower opening [
51,
52]. Exogenous auxins, indole-3-acetic acid (IAA), and naphthaleneacetic acid (NAA) can delay the DFOT of rice. The content of IAA in rice spikelets rapidly decreased within 2 h before natural opening, and the expression of IAA biosynthetic genes (
OsTAR2,
OsYUCCA3/
4/
8) in the spikelets decreased correspondingly when the spikelets opened, while the genes encoding enzymes that catalyze IAA conjugation (
OsGH3.2,
OsGH3.8), IAA efflux carrier genes (
OsPIN1,
OsPIN1a), and the gene encoding the positive regulator of the IAA polar transport factor BG1 were all significantly up-regulated [
53,
54]. This indicates that the DFOT of rice is negatively regulated by auxins.
Gibberellins are widely distributed plant hormones that have important effects on plant growth, development, flower opening, and fruiting [
7]. The “920” additive is a mixture of various homologs of gibberellins, and exogenous application of “920” can promote the flower opening of the female parent, increase the chance of overlapping DFOTs with the male parent and the hybrid seed production rate in rice [
55]. Spraying “920” before flower opening of rice sterile lines can induce an earlier DFOT by 0.5~1 h, increasing the rate of overlapping DFOTs between parents to 15–20% and improving seed production rates [
56]. Currently, “920” is widely used in rice plantation to increase the chance of overlapping DFOTs between parents and the pollination rates, and then improve seed production yields.
Triacontanol, also known as melissyl alcohol, is a natural long-chain aliphatic alcohol. As a plant growth regulator with no toxic effects, triacontanol can promote the growth and development of plant roots, stems, leaves, shoots, and flowers, and enhances various physiological functions, such as increasing the chlorophyll content and enhancing photosynthesis [
7]. Triacontanol can promote peak flower opening of sterile lines, but has almost no effect on maintainer lines in rice. Spraying the sterile line and the maintainer line at the same time with triacontanol can bring the peak flower opening times of the two lines closer and increases the rate of overlapping DFOTs [
57]. A physiological analysis shows that triacontanol can increase the activity of photosynthetic phosphorylation and promote the accumulation of adenosine triphosphate (ATP), providing energy for the flower opening of sterile lines [
58].
Huaxinling is a type of plant growth nutrient that can promote the early flower opening of parent plants and can also promote the vegetative growth and reproductive development of rice. It is a newly refined and highly effective DFOT regulator that has the effect of promoting early flower opening of the female parent and increasing the outcrossing rate [
59]. Huaxinling can increase the rate of female parent flower opening before noon by about 36.25%; it can also prolong the duration of peak flower opening of the parents, thereby partially solving the problem of non-overlapping DFOT between the parent lines [
60].