Integrins are heterodimeric transmembrane proteins that mediate adhesive connections between cells and their surroundings, including surrounding cells and the extracellular matrix (ECM). They modulate tissue mechanics and regulate intracellular signaling, including cell generation, survival, proliferation, and differentiation, and the up-regulation of integrins in tumor cells has been confirmed to be associated with tumor development, invasion, angiogenesis, metastasis, and therapeutic resistance. Thus, integrins are expected to be an effective target to improve the efficacy of tumor therapy. A variety of integrin-targeting nanodrugs have been developed to improve the distribution and penetration of drugs in tumors, thereby, improving the efficiency of clinical tumor diagnosis and treatment.
1. The Structure of Integrins
Integrins, as adhesion molecules and transmembrane receptors that consist of α and β subunits, heterodimers, are widely located on the surface of vertebrate cells and are involved in cell-cell and cell-matrix interactions [
11]. Integrins feature an extracellular domain, a single-pass transmembrane domain and a short cytoplasmic domain (except β4) [
13]. Of these, the extracellular domain can interact with ECM ligands to transmit signals [
13,
14]. Generally, there are 18α and 8β subunits that can combine to generate 24 distinct receptors with varying binding capabilities and tissue distribution [
15,
16]. Specifically, nine of these α subunits contain an αI domain at the N-terminus consisting of approximately 190–200 amino acid residues and can be strongly bound by ligands [
17]. Moreover, metal ion binding sites at the top of the αI domain have been demonstrated to bind divalent metal ions, such as magnesium ions and manganese ions, thus, mediating the interaction of integrins and ligands [
18,
19]. For the α subunits that lack the I domain (α3, α4, α5, α6, α7, α8, α9, αv, and αIIb), the β-propeller of the α subunits and the I domain of the β subunit can be joined to form the binding site [
20]. Moreover, many integrin ligands, as well as the integrins themselves, present structural alterations under specific conditions, which may compromise signaling pathways that are constitutively triggered under physiological conditions [
21,
22].
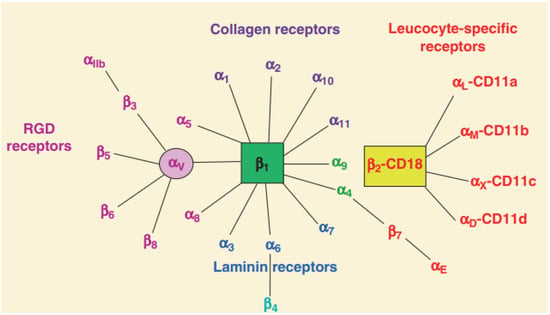
Of the 24 human integrin subtypes known to date, eight integrin dimers can recognize Arg-Gly-Asp (RGD) peptide in extracellular matrix proteins [
23]. In addition, there are four types of integrins based on their connective ligand: leukocyte, collagen-binding, Arg-Gly-Asp (RGD)-binding, and laminin-binding integrins (
Figure 1). The short cytoplasmic domain provides binding sites for these adaptors, signaling proteins, and cytoskeleton-associated proteins, all of which are required for bidirectional integrin signaling [
12,
24]. Moreover, the integrins also serve as receptors for certain toxins (such as snake venom), viruses, and other pathogens [
25]. In addition, fibronectin is regarded as a potential integrin ligand and the extra deposition of fibronectin in tissue leads to inflamed diseases [
26,
27].
Figure 1. Classification of integrins family [
8]. Integrin heterodimers were composed of different combinations of α and β subunits. In terms of ligand specificity, integrins can be divided into four groups: collagen-binding integrins (α1β1, α2β1, α10β1, and α11β1), RGD-recognizing integrins (α5β1, αVβ1, αVβ3, αVβ5, αVβ6, αVβ8, and αIIbβ3), laminin-binding integrins (α3β1, α6β1, α7β1, and α6β4), and leukocyte integrins (αLβ2, αMβ2, αXβ2, and αDβ2). The β2 integrin subunit (CD18) can pair with one of the four α subunits (αL-CD11a, αM-CD11b, αX-CD11c, and αD-CD11d), forming different leukocyte function-associated antigens.
2. The Function of Integrins
Integrins are essential for cell growth and survival. They tightly link cells to ECM and deliver signals both internally and outwardly, acting as a communication bridge between the internal and external worlds of cells [
28]. Integrins are activated in the extracellular domain via conformational rearrangement from bent-closed to open-active states [
29,
30]. In this process, the ECM ligand binding would rapidly lead to the conversion of the integrins to the high-affinity extended-open form that permits coupling to the actin cytoskeleton. This bridging to actin supports cell adhesion and the recruitment of additional intracellular binding partners that mechanically reinforce the link and allow subsequent downstream signal transduction [
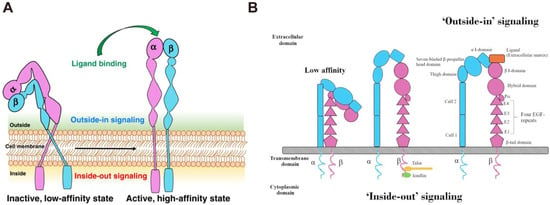
31]. The bidirectional transmembrane signals are delivered through “inside-out” and “outside-in” pathways (
Figure 2). Briefly, some intracellular signal molecules, such as talin and paxillin, can bind to the intracellular region of integrins, inducing conformational changes in the intracellular region and initiating a series of subsequent conformational changes in the extracellular domain, a process known as “inside-out” signaling [
32]. In this process, the binding of ligands to integrins causes changes in the intracellular cytoskeleton and the recruitment of important intracellular signal molecules [
33]. Furthermore, the intracellular domains of α and β subunits are separated and bind to intracellular signal molecules, activating relevant intracellular signaling pathways and mediating cell functions. This is known as “outside-in” signal transduction. The bidirectional signal transduction ability of integrins is critical for cell proliferation, invasion, angiogenesis, and apoptosis resistance, contributing to more aggressive diseases, such as tumors [
8,
34]. For example, Integrin α2 (ITGA2) is overexpressed in solid tumors, including pancreatic, gastric, liver, prostate, and breast tumor, and its expression level is highly correlated with tumor proliferation and invasion [
35,
36,
37,
38]. In addition, integrin β1, β3, β6, αv, and α5 are also found in many tumors and play an important role in tumor therapies [
39,
40,
41]. Therefore, an in-depth understanding of the interaction between integrins and tumors is beneficial in improving tumor therapy.
Figure 2. (
A) The structure and activation pathways of integrins [
8]. Integrins in the bent resting conformation reveal low-affinity binding to their ligands. After being activated, integrins were able to have a high affinity with their ligands. (
B) Schematic illustration of “Outside-in” and “Inside-out” integrins signaling pathways [
42]. Integrins can be activated from two directions, from the inside by the regulated binding of proteins to the cytoplasmic tails, and from the outside by multivalent ligand binding. For “Outside-in” signaling, signals received by other receptors foster the binding of talin and kindlin to the cytoplasmic end of the integrin β subunit, at sites of actin polymerization. As for “Outside-in” signaling, ligand binding to the external domain causes conformational changes that increase ligand affinity, modify protein-interaction sites in the cytoplasmic domains and thence the resulting signals.
3. The Effect of Integrins on Tumors
Integrins have been found to play multiple key functions in practically every stage of tumor progression, from basic tumor growth to metastasis, and hence integrin modulation has been widely used in tumor therapy [
9,
43,
44]. For example, the pre-treatment of cell monolayers with different matrix components (laminin, fibronectin, and collagen), makes neoplastic cells more resistant to cytotoxic insults mediated by different classes of chemotherapeutic agents in an event dependent on integrin-ECM components interaction [
45]. The overexpression of ECM components was proven to result in chemoresistance [
46,
47,
48]. However, because tumor incidence and growth are exceedingly complicated, many concerns remain unsolved, particularly the impact on tumor metastasis and angiogenesis [
49,
50,
51]. Further research into the mechanisms of the integrins family in tumor metastasis and tumor angiogenesis will help to expand the use of integrins in tumor therapy.
Compared with normal tissues or cells, the expression of integrins is maintained at a high level in tumors and plays an important role in tumor proliferation and invasion [
52,
53,
54]. For example, integrin β1, a member of the integrins family, has been confirmed to be overexpressed in nasopharyngeal carcinoma tissues [
37]. It promotes the migration and metastasis of nasopharyngeal carcinoma cells and reduces the sensitivity of cells to radiotherapy [
55]. It has been discovered that the inhibition of integrin β1 expression can effectively improve the radiation sensitivity of nasopharyngeal carcinoma [
56]. In addition, the high expression of integrin α5β1 in hepatocellular carcinoma is closely associated with the occurrence and metastasis of hepatocellular carcinoma, the antagonists of integrin α5β1 can significantly reduce the migration ability of hepatocellular carcinoma [
57,
58].
This entry is adapted from the peer-reviewed paper 10.3390/nano13111721