1. Vitamin D and SARS-CoV-2 Infection
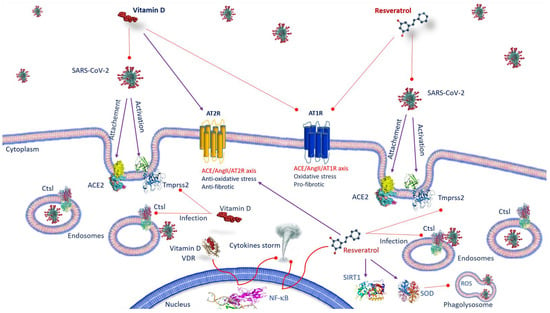
Vitamin D deficiency (VDD) is usually observed in patients developing ARDS. VDD is likely to promote pulmonary fibrosis by activating the RAAS [
28]. In considering vitamin D performance, it is important to remember the role of calcium. Low calcium intake is often associated with VDD [
29]. In coronavirus infections, calcium is important to mediate the fusion of the viral envelope with the host cell membrane or from the S1/S2 viral subunit or from the ACE2 receptor transmembrane domain [
2]. This generates the altered conversion of angiotensin II (Ang-II) to angiotensin 1–7 and the origin of a cytokine storm as well as tissue injury. Vitamin D is able to regulate the function of endothelial cells and, consequently, vasodilation. These processes facilitate ARDS onset [
2]. Thus, vitamin D safeguards from lung injury by preventing the expression of renin and the axis of the ACE/angiotensin II type I receptor (AT1R) plus stimulating the ACE2/Ang receptor (1–7)/Mas G (G-protein-Mas receptor pair) [
30]. It has been shown that, in addition, vitamin D decreases the gene expression of TMPRSS2 and Cathepsin L (Ctsl) in treated mice [
31]. Ctsl is an endosomal cysteine protease that facilitates S protein cleavage and supports viral infection through virus–host cell endosome membrane fusion [
32]. Ctsl inhibitors prevent the access of pseudotyped SARS-CoV-2 in humanized mice [
33]. Moreover, genome-wide CRISPR screening has shown that Ctsl is crucial for SARS-CoV-2 entrance [
34]. This result suggests that vitamin D impedes SARS-CoV-2 access and, thus, can moderate or prevent SARS-CoV-2 infection in lung epithelial cells [
31]. The vitamin D/VDR signaling pathway acts favorably in LPS-mediated respiratory distress syndrome by reducing the cytokine storm and by regulating the RAAS in COVID-19 patients [
35]. Moreover, it has been found that vitamin D and VDR, by the negative regulation of the RAAS cascade, and inhibition of NF-κB and Wnt/β-catenin [
36], decrease the permeability of the epithelial–endothelial barrier and counteract fibrosis in the lungs, liver, and kidneys [
36]. This action prevents the lung injury induced by sepsis, obstructing the pathway of the Angiopoietin-2-TEK receptor, tyrosine-myosin light chain kinase. Likewise, certain vitamin D metabolites are able to bind various enzymes of the SARS-CoV-2 virus replication machinery involved in the maintenance of the infection, preventing their action. Paricalcitol, a vitamin D analogue, alleviates ALI induced by LPS, preventing NF-κB activation as well as Ras expression, a member of the homologous A/Rho kinase family signaling pathways [
2]. Furthermore, vitamin D and its active hydroxyderivatives can avoid the binding of ACE2 to the receptor binding domain of SARS-CoV-2, thus avoiding SARS-CoV-2 infection. Hence, a treatment involving the integration of these compounds could be an encouraging therapeutic strategy for COVID-19 (
Figure 1) [
37].
Figure 1. Anti-SARS-CoV-2 mechanisms of action of vitamin D and resveratrol. Vitamin D, by inhibiting the binding of ACE2 to the spike protein and by decreasing TMPRSS2 and Ctsl gene expression, prevents the transmission of SARS-CoV-2. As well as vitamin D, resveratrol affects the expression of ACE-2, suppresses the effects AT1R axis, improves AT2R axis and prevents SARS-CoV-2 entry, inhibiting Ctsl and TMPRSS2. Both vitamin D/VDR complex and resveratrol inhibit NF-κB activation and cytokine storm. Resveratrol, activating SIRT1 and SOD, decreases inflammation. Abbreviations: ACE2—angiotensin-converting enzyme 2; AT1R—angiotensin II type 1 receptor; AT2R—angiotensin II type 2 receptor; Ctsl—Cathepsin L; NF-κB—nuclear factor kappa B; ROS—reactive oxygen species; SARS-CoV-2—severe respiratory syndrome coronavirus 2; SIRT1—sirtuin 1; SOD—superoxide dismutase; TMPRSS2—transmembrane serine protease 2; VDR: vitamin D receptor.
2. ACE2 Receptor and Resveratrol and SARS-CoV-2 Infection
Resveratrol affects the regulation of the RAAS and the expression of ACE-2. It suppresses the detrimental effects of the angiotensin II (Ang II)/angiotensin II type 1 (AT1R) receptor axis and improves the AT2R/angiotensin 1–7 (Ang 1–7)/Mas receptor axis, defending against renal aging [
38]. The effect of resveratrol on the two opposite pathways in the renin–angiotensin system (RAS) may equally be useful in COVID-19 patients [
20]. It has recently been found that resveratrol oligomers could prevent SARS-CoV-2 access by inhibiting Ctsl activity [
16]. Nevertheless, since Wang et al. [
16] observed that the active cavity of Ctsl is quite large for resveratrol, they suggested that resveratrol might avoid SARS-CoV-2 entry also inhibiting TMPRSS2. Then, it is fair to assume that Ctsl could be a valuable target for treating COVID-19. Resveratrol exerts its beneficial effects mainly through the activation of sirtuin 1 (SIRT1) [
39]. Stimulating both SIRT1 and superoxide dismutase (SOD), it promotes an increase in ACE2 function and a decrease in inflammation biomarkers [
40,
41]. Among the many involvements of SIRT1 in cellular processes are inflammation, stress response, and apoptosis regulation [
42]. Therefore, the upregulation of ACE2 by resveratrol may be useful in preventing SARS-CoV-2 infection and avoiding the progression of severe forms of the disease. In adipose tissue where ACE2 is significantly expressed, supplementation with resveratrol decreases both ACE2 and leptin levels, a pro-inflammatory adipokine [
43]. These effects following resveratrol supplementation could make obese individuals less susceptible to SARS-CoV-2 and could exert a beneficial impact on the outcome of the disease [
43]. On the basis of this evidence, it is worth examining the dosage range in which resveratrol has the most evident influence on the expression of ACE2, as well as the effectiveness of resveratrol supplementation on viral load and spread, and on the occurrence of COVID-19 in populations at risk, such as in obese people or those with metabolic diseases [
43]. Thus, it is important to investigate the mechanisms of action of resveratrol on the gene expression of ACE2 and the clinical outcomes of COVID-19 during infection (
Figure 1).
3. The Immune Response against SARS-CoV-2 Infection
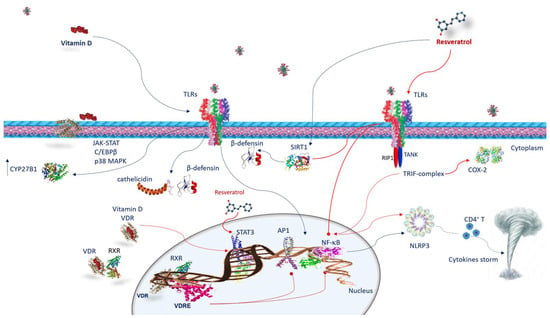
Innate immune cells recognize the pathogen-associated molecular patterns (PAMPs) and damage-associated molecular patterns (DAMPs) of virus particles through pattern recognition receptors (PRRs) such as Toll-like receptors (TLRs). Thus, identifying viral particles activates the innate immune system and triggers local inflammation liberating cytokines, IL-1β, IL-2, IL-17, G-CSF, GM-CSF, TNF-α, and chemokines such as MCP1, and IP-10, IL-8, and MIP1α (or CCL3). The cytokine storm supports a great infiltration of neutrophils and macrophages, causing severe alveolar injury [
2]. Additionally, T cells, CD4
+, Th1 and Th17, and CD8
+ T cells are the key players in the adaptive immune response, particularly in fostering the production of pathogen-specific antibodies by inducing T-dependent B cells and in the elimination of virus-infected cells. SARS-CoV-2-specific CD4
+ T cells produce IL-2, IFN-γ, and tumor necrosis factor-α (TNF-α), increasing the activation of Th1 immune response and cell-mediated immunity (
Figure 2) [
2], whereas virus-specific memory CD8
+ T cells are involved in host defense from SARS-CoV-2 infection by the secretion of IFN-γ, TNF-α, IL-2, and cytolytic molecules such as granzyme B [
44].
Figure 2. Vitamin D and resveratrol immune response against SARS-CoV-2 infection. Vitamin D interacts with the cells of the innate immunity by activating TLRs and upregulating CYP27B1. The mechanism by which TLR binding improves CYP27B1 expression involves JAK-STAT, C/EBPβ, and p38 MAPK pathways. Following TLRs stimulation, vitamin D activates innate immunity, increasing cathelicidins and β-defensins. VDR is expressed in many immune cells. Once vitamin D is hydroxylated interacts with VDR, vitamin D/VDR complex negatively regulates STAT3. Vitamin D exerts its effects through genomic mechanisms modulated by VDR/RXR complex to bind to VDRE in target genes of immune cells such as neutrophils, macrophages, dendritic cells, and T and B lymphocytes. VDRE promotes the recruitment of nuclear proteins in transcriptional-complex-modulating inflammatory response. Resveratrol acts against COVID-19 inhibiting TRIF signaling in the TLRs pathway by RIP1/TANK in the TRIF complex. It inhibits STAT3 phosphorylation. Resveratrol reduces COX-2 expression by preventing TRIF signaling. Resveratrol also activates the SIRT1 pathway, which disrupts TLR4/NF-κB/STAT signal, decreasing cytokine storm. Resveratrol-inducing SIRT1 can mediate β-defensins induction. Both vitamin D and resveratrol inhibit NF-κB and consequently, NLRP3 inflammasome, avoiding CD4+ T cell activation and cytokine storm. Abbreviations: AP1—activator protein 1; C/EBPβ—CCAAT/enhancer binding protein β; CD4T—CYP27B1, cytochrome P450 family 27 subfamily B member 1; COX2—cyclooxygenase-2; JAK-STAT—Janus kinase/signal transducer and activator of transcription; NF-κB—nuclear factor kappa B; NLRP3—NLR family pyrin-domain-containing 3; p38 MAPK—p38 mitogen-activated protein kinase; RIP1—receptor-interacting protein; RXR—retinoid X receptor; SIRT1—sirtuin 1; STAT 3—signal transducer and activator of transcription 3; TANK—TRAF family member-associated NFKB activator; TLR—Toll-like receptor; TRIF—TIR-domain-containing, adapter-inducing interferon-β; VDR—vitamin D receptor; VDRE—vitamin D response elements.
4. Anti-Inflammatory Activity and Immune Response of Vitamin D and Resveratrol against SARS-CoV-2 Infection
Both vitamin D and resveratrol, constraining the secretion of pro-inflammatory cytokines, exert anti-inflammatory activity, and play an immunomodulatory role in both adaptive and innate immune systems [
2,
17,
45] (
Figure 2). Notably, cells of the immune system such as neutrophils, monocytes, macrophages, dendritic cells, B cells CD4
+, and CD8
+ T cells express VDR; hence, they are targets of vitamin D. Additionally, most of them, through VDR, are able to activate and inactivate vitamin D metabolites [
45]. Likewise, resveratrol activates macrophages, T cells, and natural killer (NK) cells, and is involved in the CD4
+CD25
+ regulatory T cell immunosuppressive role [
19].
4.1. Vitamin D and Innate Immune Response against SARS-CoV-2 Infection
The actions of vitamin D on the immune system are largely determined by the vitamin D receptor (VDR). VDR belongs to the family of nuclear steroid receptors and is expressed in tissues of the whole body, including many immune cells [
2], and affects many cellular functions. VDR operates by binding with its ligand [1α,25(OH)2D3] and creating a heterodimer with the retinoid X receptor (RXR). The VDR/RXR complex, by binding the proximal-promoter VDR response element (VDRE), promotes the enrollment of nuclear proteins in the transcriptional complex and modulates gene expression. The interplay of vitamin D with the cells of the innate immune system occurs through the activation of TLRs. It has been found that VDR-binding 1,25(OH)
2D
3 can promote TLR2/1 heterodimer ligation in macrophages, upregulating CYP27B1 [
2]. The CYP27B1 gene provides instructions to generate an enzyme called 1α-hydroxylase. Human 1α-hydroxylase promoter analysis has identified binding sites for various transcription factors such as NF-κB, AP1, AP2 and Sp1; cAMP binding sites (CRE); a CCAAT box; and a GAS.3 site [
46]. TLR binding enhances CYP27B1 expression, involving JAK-STAT, C/EBPβ, and p38 MAPK pathways [
47]. The activities of these factors can be controlled by inflammatory stimuli. Several TLRs, such as TLR2, TLR3, TLR4, TLR6, TLR7, TLR8, and TLR9 are potential targets in controlling the early stages of COVID-19 infection [
48]. It has been shown that vitamin D induces TLR2 and TLR4 [
2,
45]. Moreover, vitamin D regulates TLR9-dependent IL-6 production and increases TLR7 and NE/PAD4/COX-3/GAPDH [
49]. In human macrophages, as a result of TLR2/1 stimulation, vitamin D activates innate immunity increasing the antimicrobial peptide levels of cathelicidins and β-defensins (
Figure 2) [
17]. These findings strongly suggest that individuals with sufficient vitamin D levels are able to counteract TLRs and subsequent AMPs and have adequate cytokine production [
17].
4.2. Vitamin D and Adaptive Immune Response against SARS-CoV-2 Infection
Vitamin D powerfully influences macrophage polarization. In particular, it blocks the polarization from the M1 phenotype towards the M2 by decreasing IFN-γ release and promoting IL10 production [
2]. The M2 phenotype inhibits inflammation, stimulating type 2/Th2 immune responses and producing IL-10, which in turn, prevents the M1 polarization. [
2]. This is also fundamental in the conversion of pro-inflammatory Th1 cells versus regulatory T cells (Tregs) to resolve inflammation in severe COVID-19 through the production of TGF-β, CCL22 and additional IL-10 [
2]. Therefore, the production of vitamin D by macrophages results in a modification from an inflammatory to a tolerogenic state [
2]. Interestingly, vitamin D, increasing IL-4, IL-5, and IL-10 and reducing Th17 cells and IL-17 production, upregulates the Th2-specific transcription factors, GATA-3 and c-maf. In addition, IL-17 reduction inhibits the inflammatory response dominated by neutrophils. Th17 cell proliferation is induced by signals mediated by IL-6 IL-21, IL-23, TGF-β, and by the retinoic-acid-related orphan nuclear receptor (RORγT), which is a lineage-specifying transcription factor. Other crucial targets of vitamin D are dendritic cells (DCs) that belong to antigen-presenting cells (APCs), which affect lymphocyte activation, prompting the adaptive immune response and maintaining peripheral tolerance (
Figure 2) [
2]. Remarkably, the vitamin D–VDR complex binds to the promoter of STAT3, IL-6, and IL-10 genes. Furthermore, vitamin D, influencing T cell responses, indirectly acts on B cells. Since B cells express VDR and CYP27B1, these cells are also predisposed to vitamin D effects, including the inhibition of their proliferation and plasma cell differentiation, and the induction of apoptosis. Consequently, vitamin D is capable of modulating antibody production [
2].
4.3. Resveratrol and Innate Immune Response against SARS-CoV-2 Infection
Recent evidence has demonstrated that resveratrol strongly prevents viral replication of both DNA and RNA viruses [
50,
51]. An in vitro investigation also demonstrated that resveratrol decreases viral replication of SARS-CoV-2 and prevents cytotoxicity [
50]. A randomized, double-blind clinical trial confirmed that resveratrol can efficiently serve as a treatment adjunct for COVID-19 [
52]. In silico examinations reported that resveratrol is able to bind to the ACE2 complex [
53]. Resveratrol is also able to decrease the effect of proinflammatory cytokines such as IFN-γ, TNF-α, and IL-1β, which are important factors in the onset of the cytokine storm in COVID-19 [
20]. The anti-inflammatory action of resveratrol has been linked to the suppression of NF-κB activation [
54] by facilitating the inhibition of Iκ-B kinase [
55]. The NF-κB pathway is strongly associated with the pathogenesis of numerous inflammatory diseases and the manifestation of severe inflammatory sequelae [
56]. It is well known that an exaggerated activation of NF-κB elicits the evolution of the inflammatory response with the production of many proinflammatory mediators such as IL-1, IL,6, TNF-α, IFN-γ, interleukin-8 (IL-8), granulocyte-macrophage-colony-stimulating factor (GM-CSF), COX-2, and inducible nitric oxide synthase (iNOS) [
57]. Numerous immunosuppressive or anti-inflammatory agents usually target the NF-κB pathway [
58]. In addition, NF-κB stimulates the activation of the NLR family pyrin domain containing the 3 (NLRP3) inflammasome by facilitating the priming signal of the inflammasome. The NLRP3 inflammasome induces the maturation and secretion of pro-inflammatory cytokines, ultimately promoting CD4
+ T cell differentiation and activation [
59]. Therefore, the inhibitory effect of resveratrol on NF-κB results in the decline of the expression of inflammation-associated genes, supplying a suitable target for anti-inflammatory treatments (
Figure 2) [
17]. It has been found that resveratrol reduces COX-2 expression by preventing TIR-domain-containing, adapter-inducing interferon-β (TRIF) signaling associated with TLR3 and the TLR4 MyD88-independent pathway through TRAF family member-associated NF-κB activator (TANK)-binding kinase 1 and receptor-interacting protein 1 (RIP1) in the TRIF complex [
17]. Thus, it is possible that the action of resveratrol against SARS-CoV-2 occurs through specific inhibition of TRIF signaling in the TLR3 and TLR4 pathways by involving kinase 1 and the RIP1 TANK binder in the TRIF complex. However, resveratrol activates other anti-inflammatory pathways, including Sirt1 [
17]. This latter inhibits the TLR4/NF-κB/STAT signal, which, in turn, diminishes cytokine release from inactivated immune cells [
17], or macrophage/mast-cell-derived pro-inflammatory factors, as the platelet-activating factor, histamine and TNF-α [
60]. Other targets of resveratrol are pathways influencing immune function and the cellular inflammation response. For example, it can cause the conversion of cellular metabolism and signal transduction pathways by distressing enzymes, such as adenosine monophosphate kinase and the serine/threonine protein kinase that is the mechanistic target of rapamycin (mTOR) [
58]. Overall, the antiviral mechanisms of resveratrol versus SARS-CoV-2 can act through the inhibition of virus replication, synthesis of proteins, and inhibition of transcription and signaling pathways, along with viral-related gene expressions [
17]. Some evidence has indicated that resveratrol, in response to microbial agents such as
Haemophilus influenzae and
Streptococcus pneumoniae [
61,
62], is capable of inducing the production of several classes of antimicrobial peptides (AMPs) such as α- and β-defensins and cathelicidin [
63]. Therefore, it is conceivable that resveratrol and its analogues, through SIRT1 protein, also mediate β-defensin induction following SARS-CoV-2 infection. Resveratrol, reestablishing glutathione concentrations, can interfere with the differentiation of monocytes to macrophages and inflammation (
Figure 2) [
64]. Moreover, if on the one hand, resveratrol regulates the M2 macrophage activation and differentiation impeding IL-10 and MCP-1 productions, on the other, it promotes TGF β1 production [
17]. Nevertheless, resveratrol inhibits the phosphorylation of STAT3 without interfering with its expression in the M2 macrophage differentiation.
5. Vitamin D and Resveratrol Effects on Neutrophil Extracellular Traps (NETS) in SARS-CoV-2 Infection
In severe COVID-19 patients, the number of neutrophils increases [
65]. Neutrophils are phagocytic cells that, with their microbicide activity, their degranulation mechanisms, and their release of neutrophil extracellular traps (NETs), play an essential role in the defense against infectious diseases. NETs are web-like chromatin structures consisting of cytosolic and granule proteins that are collected on a support of uncondensed chromatin. NETs originate from the nucleus but also contain mitochondrial DNA [
66]. NETs contain abundant enzymatic proteins such as neutrophil elastase, myeloperoxidase, and peptidyl arginine deiminase 4. Therefore, NETs have antibacterial activity able to eradicate any kind of microbial agent [
66]. Some COVID-19 patients have exhibited elevated levels of NETs [
67]. Therefore, their dysregulation is implicated in the COVID-19 pathogenesis. NETs promote cytokine release, initiating a signal cascade of the inflammatory type that causes micro-thromboses and pulmonary, cardiovascular, and renal system complications [
68]. It has been observed that vitamin D can control the proliferation of pathogens, such as viruses, by inducing NETs [
69]. However, the role played in the initiation of NETs by vitamin D in SARS-CoV-2 appears to be ambivalent [
2,
70]. In contrast, it has been found that resveratrol reduces NET arrangement in severe COVID-19 patients mitigating neutrophil activation [
71].
This entry is adapted from the peer-reviewed paper 10.3390/nu15112639