Interest in ExP-producing bacteria has also expanded into the research areas of environmental sciences, including studies on eco-friendly municipal and wastewater treatment processes [
3]. This is because many standard remediation techniques require the usage of reagents that may be hard to degrade, or various environmentally harmful by-products are produced during their utilization at contaminated sites. This includes chemical treatment, which usually detoxifies metals via redox transformation or neutralization by application of the reagents, such as potassium permanganate, hydrogen peroxide, hypochlorite, synthetic surfactants, or chlorine gas, to precipitate, immobilize, or preconcentrate the hazardous contaminants [
4,
5,
6]. The chemical leaching of soils and sediments by applying strong inorganic and organic acids and persistent synthetic chelating agents (e.g., ethylenediaminetetraacetic acid or its derivatives) to solubilize contaminants has also been successfully tested for heavy metal removal [
7]. Reactive solid inorganic and biological substances, as well as materials with active surfaces (e.g., zero-valent iron, ferric oxides and oxohydroxides, nanomaterials, zeolite, biological waste) have been studied as potential components of permeable treatment barriers to restrict the movement of the contaminant in the environment [
8,
9,
10,
11]. However, in some cases, unpredictable effects regarding the toxicity and mobility of generated species can be expected since these interactions are usually non-specific. The application of electrochemical and electrokinetic remediation methods, engineered to site-specific requirements, has been performed for heavy metal removal [
12], showing promising results in combination with other remediation approaches, including novel biochemical methods [
13]. More prominent green approaches include phytoremediation, phytoextraction, and biosorption. They are usually performed in conjunction with other methods, e.g., chemical leaching [
14]. Still, the application of microbial ExPs is environmentally advantageous since these biogenic polymers are usually water soluble, susceptible to natural degradation, and less harmful than synthetic polymers [
15].
Bacterial ExPs find their successful applications in heavy metal removal, oil recovery, and various in situ remediation techniques such as emulsifiers, sorbents, biofilters, surfactants, and bioflocculants [
16,
17,
18]. The interest of environmental researchers in ExP-producing bacteria is also highlighted in several patent deposits focusing on the prosperous application of bacteria in the remediation of contaminated sites. Villela et al. [
19] reported that there are 114 patents describing the degradation of oil compounds exclusively by
Pseudomonas, thus highlighting the leading role of this bacterial genus in hydrocarbon-contaminated site remediation.
The utilization of
Pseudomonas strains in remediation is not limited solely to the biodegradation of hydrocarbons; they have also been successfully applied for the decontamination of heavy metal-polluted waters, soils, and sediments [
20]. This is primarily due to their ability to produce metal-chelating siderophores and surface-active extracellular polymeric substances [
21]. Regarding the latter,
Pseudomonas species are considered high-ExP-producing organisms [
22], and since they are ubiquitous, being isolated from various types of environments, including industrial waste and activated sludge [
23], they are considered potent in solving the issue of heavy metal contamination [
24].
2. Biosynthesis of Extracellular Polysaccharides in Pseudomonas
Extensive progress has been made in elucidating the synthesis of bacterial extracellular homopolysaccharides and heteropolysaccharides in recent years (
Figure 1). They are synthesized by bacteria either extracellularly (outside the cell membrane and the cell wall), within the cell wall, or intracellularly [
25].
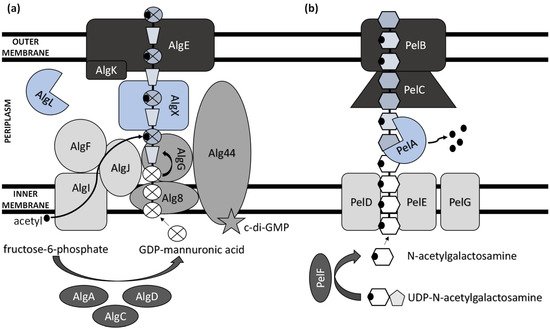
Figure 1. Schematic model of (a) alginate and (b) Pel polysaccharide biosynthetic machinery.
While heteropolysaccharides are mostly synthesized intracellularly and transported outside the cell, homopolysaccharide production generally involves the activity of enzymes secreted by the bacterium to the extracellular environment. ExP production comprises several steps, including the synthesis of ExP precursors, repeat-unit assembly on a lipid carrier located at the cytoplasmic membrane, modification (e.g., acylation, acetylation, sulphation, and methylation), membrane translocation, polymerization, and export [
26]. Thus, there are several functionally distinguished enzymes required for ExPs’ synthesis and development [
27]. Several other enzymes, which are not unique to ExP production and serve as intermediates in protein regulation and central carbon metabolism, are also involved in ExPs’ biosynthesis process [
28].
The biosynthesis of ExPs requires the involvement of activated monosaccharides derived from catabolized sugars. These include sugar nucleosides (nucleoside diphosphate sugars) or their derivatives (e.g., uridine diphosphate (UDP)-N-acetylglucosamine and guanosine diphosphate (GDP)-mannuronic acid) [
29].
The main ExP component in the bacteria of
Pseudomonas genera is alginate, an anionic linear polymer composed of beta-1,4-linked mannuronic acids (M-blocks) and C5-epimer α-L-guluronic acid (G-blocks) [
30]. Alginate’s viscosifying, gelling, and stabilizing properties make this biopolymer an important industrial polysaccharide with great prospects in applications such as drug and protein delivery systems and food encapsulation [
31,
32]. Thirteen proteins are directly involved in the biosynthesis of alginate, and except for AlgC, they are all encoded by the
alg operon [
33]. The alginate precursor is synthesized by three enzymes including AlgA (bifunctional enzyme phosphomannose isomerase/guanosine 5′-diphospho-D-mannose pyrophosphorylase), AlgC (phosphomannomutase) and AlgD (GDP-mannose dehydrogenase), which allow the conversion of fructose-6-phosphate to GDP-mannuronic acid via four steps [
34,
35,
36]. Alginate is first synthesized as a linear homopolymer from the GDP-mannuronic acid to polymannuronic acid by catalytic subunit Alg8 (alpha-1,3-glucosyltransferase) which interacts with Alg44 co-polymerase located at the cytoplasmatic membrane [
37], with most of the latter being exposed to the periplasm [
38]. These enzymes allowed the movement of the alginate precursor across the inner membrane for polymerization [
39]. Alg44 also demonstrated the capability of binding the second messenger cyclic dimeric guanosine monophosphate (c-di-GMP) synthesized by MucR, a membrane-anchored protein, which is required for alginate biosynthesis [
40].
In the periplasm, the polymannuronate is modified by epimerization or acetylation. AlgI, AlgJ, and AlgF are required for the addition of O-acetyl groups to the alginate polymer at O2 and/or O3 positions, which is an essential process for the stabilization of the intracellular alginate matrix for microcolony formation [
41]. Acetylation can also affect epimerization reactions, since the activity of AlgG, a C5-epimerase that directly converts D-mannuronate to L-guluronate, has been detected only when the acetyl groups are removed from the polymannuronate substrate [
42]. Newly formed macromolecules are most likely transported within the periplasm by the periplasmic protein AlgX that surrounds and protects the polymers from degradation by AlgL, a periplasmic alginate lyase [
43,
44]. Alginate is then secreted by the putative export protein AlgE [
45]. The proper localization of AlgE for the periplasmic components of the alginate’s biosynthetic machinery is facilitated by AlgK [
46].
The acid hydrolysis method is usually applied for the determination of ExPs’ monomeric components (e.g., glucose, fructose, galactose, and arabinose). Myszka and Czaczyk [
47] reported that, under starvation conditions (ABPG medium reduced by 90% (
w/
v) of optimal nutrient availability), the EPS matrix of
Pseudomonas aeruginosa ATCC 10145 consisted solely of glucosyl units. Grob, et al. [
48] suggested that the survival of
P. aeruginosa SG41 under highly chlorinated conditions was enabled by the overproduction of alginate, a major component of the SG41 strain’s ExPs (109.8 µg·g
−1 cell dry mass) [
49]. Thus, alginate overproduction is advantageous in harsh environments. Still, the nonmucoid
P. aeruginosa strains that are the predominant environmental phenotype do not need to express the alginate biosynthetic genes to form the nonmucoid biofilms [
50]. These use either Pel or Psl as the primary matrix structural polysaccharide [
51].
A previously reported analysis of Pel polymer suggested that it is rich in cationic amino sugars, N-acetylgalactosamine, and N-acetylglucosamine, in a 5:1 ratio [
52]. However, just recently, Le Mauff et al. [
53] characterized the configuration and structure of Pel and suggested that it is a polymer of partially de-N-acetylated α-1,4-N-acetylgalactosamine, and it does not contain N-acetylglucosamine.
Pel synthesis requires protein products of a seven-gene operon
pelABCDEFG [
54]. The protein complex of PelD, PelE, PelF, and PelG is very likely a Pel-polysaccharide synthase whose activity is dependent on the localization of cytosolic glycosyltransferase PelF [
55] to the inner membrane protein complex PelDEG [
56]. PelDEG is also competent for the transport of the Pel polymer across the cytoplasmatic membrane.
PelA is a multi-domain protein that localizes to both the periplasm and membrane and exhibits both hydrolase and de-N-acetylase activity [
57,
58]. The PelBC complex is responsible for the transport of the matured polymer into the extracellular milieu. PelB is located at the outer membrane and contains a transmembrane β-barrel porin towards which the lipoprotein PelC, which is localized to the inner leaflet of the outer membrane, guides the positively charged Pel and, thus, acts as a charged molecular funnel facilitating Pel export [
59].
Psl is a neutral branched pentasaccharide comprising D-mannose, D-glucose, and L-rhamnose. It is synthesized by the
polysaccharide
synthesis
locus (
psl). The
psl gene cluster consists of 15 genes, of which 11 are necessary for Psl polysaccharide synthesis (
pslACDEFGHIJKL) [
60]. However, the specific function of each protein is not completely understood.
pslB likely encodes a bifunctional enzyme with GDP-mannose pyrophosphorylase/phosphomannose isomerase dual activities, which is the only enzyme from the
psl operon involved in sugar nucleotide precursor production [
61]. The inner membrane-associated glycoside hydrolase PslG can be involved in the biosynthesis of Psl polysaccharide [
60], although its role in this process is controversial [
62]. Since the five PslAEJKL proteins have inner membrane-spanning domains, it was hypothesized that they make up the Psl polymerization complex [
33]. Regarding Psl translocation and export, the complex of PslD and PslE helps transport Psl across the outer membrane [
63].