1. Introduction
Every individual in an animal population differs from the other members in morphology, physiology, and behavior. Much of this diversity is due to genetic variability in the members of a population, i.e., subtle genetic differences between the individuals [
1,
2,
3,
4]. Genetic variability in animal populations is the prerequisite for evolutionary processes [
5]. However, even in clonal organisms, which are genetically fully identical, differences in trait patterns occur; these are products of either coincidence [
6,
7] or differences in the environmental conditions during the embryonic development or other periods of an animal’s life span [
8]. Furthermore, alterations in environmental conditions during the adult life of an animal may affect the structure and function of its cells and organs [
9]. ‘Phenotypic plasticity’ is the ability of an animal to adjust the individual phenotype in response to certain stimuli from the environment or from endogenous sources in the absence of any changes in its genome [
10,
11]. Examples of this include the processes of skin tanning by sunlight (environmentally induced plasticity) or the increase in muscle mass in response to muscular work in humans (functionally induced plasticity).
In most cases, phenotypic plasticity is of adaptive value and increases the individual’s fitness under unstable environmental conditions. In such cases, the bandwidth of phenotypic plasticity with respect to a given trait is genetically defined and is itself subject to selection [
12,
13,
14,
15,
16,
17,
18,
19,
20,
21]. However, in some cases the environmentally induced phenotypic changes may be merely passive consequences of changes in environmental parameters (e.g., reduced growth rate when food is scarce and the animals are undernourished). In such cases, there is no adaptive value of phenotypic changes resulting from environmental change [
22,
23,
24,
25,
26]. To judge the ecological significance of observed cases of phenotypic plasticity, it is necessary to determine whether individual fitness is maintained or increased (beneficial acclimation hypothesis) or not (detrimental acclimation hypothesis) [
27]. The current ecological models consider these possibilities [
28,
29].
2. Measure for Plasticity in Response to Environmental Change: The Reaction Norm
Most animal species are able to survive and produce offspring in environments which undergo changes in various abiotic and biotic parameters (salinity or pH of the surrounding medium, environmental temperature, food availability, etc.). The course and the extent to which a certain trait is changed in response to such shifts in environmental conditions are described by ‘reaction norms’ [
1,
30,
31]. Comparisons of reaction norms enable ecologists to quantitatively describe imminent changes in life cycle parameters in different animals (body size, motility, fertility, etc.) and their dependency on defined shifts in environmental conditions (
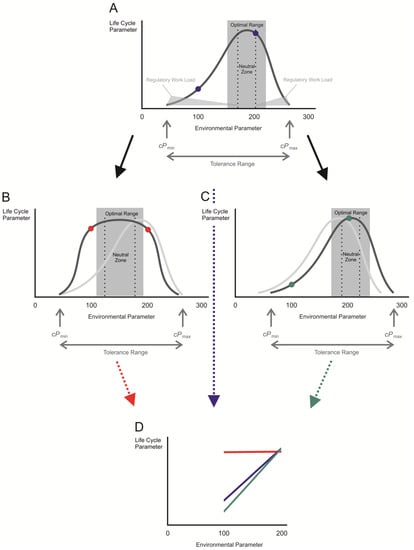
Figure 1).
Figure 1. Dependence of a life cycle parameter in an animal on the condition of its environment and the potential adjustments during acclimation. The original tolerance curve (A) may change in width (B) or position on the x-axis (C) (original is a grey shadow for comparison) during acclimation processes. Such changes result in alterations in the slopes of reaction norms (D) as exemplified by comparing the resulting reaction norms within the limits of the colored spots in A, B, and C, which represent the same pair of environmental conditions. cPmin—critical minimum of the environmental parameter; cPmax—critical maximum of the environmental parameter. Further explanations in the text.
There is a strong dependence of virtually every life cycle parameter in an individual animal on the exact value of a given environmental condition. The actual values of this life cycle parameter over the range of tolerable environmental conditions are represented by an optimum curve, the ‘tolerance curve’. The ‘optimal range’ is the interval of values around the maximum of the tolerance curve. In this range, the animal needs to divert only a little energy or no energy from other bodily functions towards maintaining its internal conditions, while keeping the life cycle parameter of interest at its maximum. Within the optimal range, the ‘neutral zone’ marks an interval of values of the external conditions at which the organism is in equilibrium with its environment. Under these conditions, the internal parameters do not to have to be defended against the external ones due to a lack in the gradients between the internal space and the environment or in disturbances of the physiological conditions within the body. The amount of energy that has to be invested to keep internal homeostasis, the regulatory workload, is at its minimum within this neutral zone (Figure 1).
However, if the environmental conditions lie outside the optimal range, increasing amounts of work have to be invested in homeostatic functions. The energy required for this purpose is then diverted away from the sustaining of the life cycle parameter of interest so that its value becomes smaller as we approach the upper or lower critical limits of the environmental condition (cPmax or cPmin, respectively). Beyond these limits, the amount of energy required for maintaining internal homeostasis would become so large that sustained life would become impossible. These limits define the ‘tolerance range’ of an individual.
Theoretically, the reaction norm of an animal for a given life cycle parameter (e.g., the growth rate) is defined by two or several points within the tolerance curve. Using only two points of different environmental parameter values results in a linear reaction norm with a certain slope (Figure 1A,D, blue dots or line, respectively). This type of reaction norm is only valid for a given interval of the environmental parameter and may be used to compare the behavior of different individuals from the same or from different populations in their responses to this particular environmental change. However, the linear reaction norm derived from the two points on the tolerance curve provides only simplified information on the response of an animal to environmental change and may miss important alterations in life cycle parameters when the interval of the environmental parameters has been selected in an unfavorable manner. The determination of a reaction norm using several different values of environmental parameters within the tolerance range is more informative. The best way of understanding the responsive behavior of an animal to environmental change is by the construction of a non-linear reaction norm which forms an approximation to the tolerance curve. This requires measurements of different values of the life cycle parameter of interest in small intervals of the environmental parameter within the tolerance range; thus, a lot more experimental work is needed.
An animal existing under environmental conditions outside of the optimal range but still within the critical limits experiences ‘physiological stress’ [
32]. The regulatory work that this animal has to invest to stay alive is larger than that required when the animal’s environmental condition remains within the optimal range. If energetic resources are not limited, the animal may live under such unfavorable conditions for a long time. It is noteworthy that the responses of an animal living under the stress conditions represented by the left portion of the tolerance curve are entirely different from those that are required if the animal is exposed to conditions represented by the right portion of the curve.
As energy is usually a limiting factor in an animal’s life, it is unlikely that an animal will maintain high metabolic rates for sustained periods to cope with physiological stress. Instead, the animal will activate gene regulatory, biochemical, or physiological mechanisms [
33] to reduce metabolic demands, to protect organs and tissues from damage, or to optimize the functions of regulatory organs to match the increasing regulatory demands [
34,
35,
36,
37,
38]. These environmentally induced measures in an animal are summed up as ‘acclimation’ (in cases when animals adjust to new environmental conditions induced by an experimenter in the laboratory) or as ‘acclimatization’ (when wild animals adjust to changes in their natural environments) [
25,
39].
3. Acclimation
The exposure of an animal to physiological stress generally alters both the tolerance limits and the reaction norms for the relevant physiological traits. During acclimation, the tolerance curve may be broadened (Figure 1B) or shifted on the x-axis (Figure 1C). Such acclimation processes may result in changes in the position and/or the slope of the reaction norm of relevant physiological or other life cycle parameters (Figure 1D) when those of naïve (Figure 1A) or of acclimated animals are compared (Figure 1B,C).
Environmentally induced alterations in gene transcription, in the translation rates of certain transcripts, in the posttranslational modifications of proteins, or in the epigenetic mechanisms may be involved in such responses of animals to environmental stress [
33,
40,
41,
42]. This usually requires the sensing of the new environmental condition, the systemic processing of information [
34,
43,
44,
45,
46,
47,
48,
49], and, at the cellular level, the initiation of signal transduction and activation of transcription factors [
50]. In many cases, processes such as cell differentiation or cell proliferation in tissues or organs are required to enable an animal to successfully cope with the stressful conditions [
51].
The experience of stressful environmental conditions may not only result in acute changes but may also create an elevated level of resilience which enables animals to respond quicker or more economically to further periods of environmental stress once they have successfully dealt with such stress conditions before. These ‘hormetic’ effects, which may last for different periods of time (’memory´), may play important roles in hardening animals against further periods of environmental stress [
52].
Animal physiologists use the term ‘physiological adaptation’ to differentiate this type of adjustment in single individuals from ‘genetic adaptation’, which is an evolutionary process involving several generations of animals, genetic recombination, and selection. Ecologists, however, avoid the term ‘adaptation’ when discussing environmentally induced alterations in individual animals and prefer to use the term ‘adaptive phenotypic plasticity’ [
53,
54]. When such plastic processes in response to changing environmental conditions occur during ontogenesis (e.g., during embryonic development), the alterations in the phenotype are subsumed under the term ‘developmental plasticity’ [
1,
55,
56].
4. Reversibility of Plastic Changes
Whether adaptive phenotypic changes are reversible or not depends on the biological limitations of the respective organ systems in the animals and on the nature of the environmental condition. The organ system that responds to environmental change must have the ability to respond in different directions, and the direction should be correlated with the actual environmental alteration. However, the patterning of a certain type of environmental alteration (frequent or infrequent events) also affects the kind of response in the animal.
Chronic changes in an environmental condition or the expectation that an acute change may prevail or at least occur frequently in the future may trigger irreversible changes in cell and organ functions that are maintained for the rest of the animal’s lifetime. Such cases represent ‘phenotypic plasticity in the narrow sense’.
However, there are also cases in which environmentally induced plastic changes are reversible once the environmental conditions have returned to the original ones. In such cases, the plastic changes may be induced and reversed many times during the remaining lifespan of an animal. Examples include the seasonal changes occurring in inhabitants of the temperate zones [
57], e.g., in hibernators [
58] or in animals that change fur thickness in preparation for summer or winter, respectively. When the environmental alterations occur in such a way that the animal is affected unexpectedly or if transient changes in such conditions occur from time to time without a clear rhythm (fluctuation) or take place only rarely throughout an animal´s lifetime, it is especially important for an animal to be able to respond to such alterations in a reversible manner. In such cases, it is more economical for an organism to react acutely by investing in molecular and cellular reorganization or organ optimization and to reverse these processes as soon as their effects are not needed any more, rather than maintaining the altered phenotype even when conditions have normalized again. This helps to save energy which would otherwise have to be spent on the maintenance of obsolete traits for long periods.
Some authors call the ability of organisms to reverse phenotypic changes in response to environmental stimuli ‘phenotypic flexibility’ [
6,
59,
60,
61]. However, this term is rather general and does not reflect the fact that it needs biological work and energy to reach a new steady state in an altered environment and to reverse it after the environmental alteration which had triggered the response is terminated. Thus, researchers suggest that the term ‘phenotypic elasticity’ is better suited for describing reversible adjustments in animals to changes in environmental conditions. This term allows the clear differentiation between reversible alterations of traits in animals to transient changes in the environmental conditions and those that can only be induced once in the lifetime of an organism (‘phenotypic plasticity in the narrow sense’) [
62]. In addition, this term allows the differentiation between phenotypic changes that are advantageous (albeit energetically costly) for an individual (and thus are of adaptive value for the organism) and all others.
This entry is adapted from the peer-reviewed paper 10.3390/physiologia3020025