1. Mycorrhizal vs. Non-Mycorrhizal Plants
For relevant information regarding classification of mycorrhizal associations, as well as characterization of functional aspects of mycorrhizal symbiosis, readers are invited to consult recent articles [
29,
30,
31]. It needs to be mentioned, however, that, in grassland habitats, arbuscular mycorrhizal symbiosis is of primary importance, but there are also other types of mycorrhiza present, such as orchid mycorrhiza.
Most importantly, in order to understand the importance of mycorrhizal symbiosis, we should first determine whether mycorrhizal symbiosis is a widespread phenomenon in coastal grasslands. On the one hand, abundance, frequency, and anatomical diversity in mycorrhizal associations have been assessed in different coastal habitats, including grasslands [
32]. On the other hand, a number of studies show the importance of mycorrhizae in the adaptation of plants to environmental factors, which are characteristic of coastal grasslands. For example, there is a considerable amount of research on the importance of mycorrhizal symbiosis in halophytes and its potential to increase salt tolerance in glycophytes. Interestingly, the first observations on mycorrhizal symbiosis in wild plants were from studies in coastal salt marshes and included several halophytic species [
33].
One of the problems in assessing the functional importance of mycorrhizae in coastal grasslands is related to the limited universal nature of mycorrhizal symbiosis. This manifests as a low mycorrhizal intensity in different situations or even the appearance of non-mycorrhizal plant taxa. It needs to be emphasized that it is generally accepted that a relatively low degree of root mycorrhization does not automatically mean little functional importance, and, since the intensity of symbiosis is a highly variable quantity, it is easy to overlook the situation when individuals of a given plant species show significant signs of mycorrhization. Thus, even species described as non-mycorrhizal, such as
Triglochin maritima, showed mycorrhizal structures in roots, such as vesicles and arbuscules, suggesting the presence of functionally active symbiosis but with relatively low intensity [
32]. However, this species did not show any signs of mycorrhizal colonization in the previous studies [
34], and genus
Triglochin has been considered to be non-mycorrhizal [
35]. Seasonal changes in mycorrhizal colonization showed that, in roots of
Triglochin maritima, plants’ intensity of mycorrhizal symbiosis increased from less than 5% in May to 25% in July, but the presence of arbuscules was very low in May and June but increased to 15% in July, reaching 25% in September [
11]. Therefore, it can be hypothesized that the two dominant environmental factors are associated with the low intensity of mycorrhizal symbiosis on plants in relatively moist coastal grasslands, namely high salinity and soil flooding. These aspects will be analyzed further.
Given the fact that several plant families with significant halophyte occurrence (Ateraceae, Brassicaceae, Chenopodiaceae, Carophyllaceae) have been reported as essentially mostly non-mycorrhizal [
35], the question of the general importance of mycorrhizal symbiosis in salt-affected habitats remains open.
2. Mycorrhizal Fungal Community Structure
Assessment of genetic diversity in arbuscular-mycorrhiza-forming fungal communities is an important aspect of microbial ecology. Because mycorrhizal fungi are associated with particular plant species and are subject to seasonality and environmental conditions, both the diversity and occurrence of mycorrhizal fungi are highly variable. Ecosystem-level comparison of different studies on the community structure has revealed that some Glomeromycota taxa are found globally, while others can be found only in certain ecosystems [
36]. Ecological aspects of arbuscular mycorrhizal symbiosis in halophytic plant species have been reviewed, and it has been suggested that diversity in mycorrhiza-forming fungi seems to be more complex than usually assumed [
37].
Only some studies so far have addressed community structure of mycorrhizal fungi in coastal grasslands. Using mycorrhizal roots of a single common plant species,
Hieracium pilosella, high spatial diversity in fungal phylotypes was found in a sandy coastal grassland [
38]. However, even a single root fragment from an individual plant contained almost all genetic variation found within the whole area. Most importantly, it was shown that there is a possibility that a single individual non-sporulating mycelium might cover an area at least 10 m in length. In another study, it was tested if the dominant mycorrhizal fungal strains found in roots of
Hieracium pilosella can colonize individuals of other abundant plant species,
Hypochaeris radicata,
Thymus serphyllum,
Artemisia campestris, and
Armeria maritima [
39]. As was expected, the dominating strains were found in root fragments of all five plant species but with spatial differences in intensity of occurrence. Therefore, it was concluded that presence of dominant fungal strain is an indication of presence of interconnecting mycelial mycorrhizal network in a coastal grassland.
The effect of changes in various environmental factors on the structure of mycorrhizal fungal communities in coastal grasslands has been studied very little. It should be assumed that the heterogeneity of dominant factors significantly affects this structure. Thus, changes in community structure of AM fungi have been assessed in respect to reclamation of saline coastal lands, and it was shown that vegetation succession following reclamation results in a decrease in overall fungal diversity and a shift from dominance of Acaulosporaceae and Gigasporaceae to Glomeraceae [
40]. In salt-affected coastal plains, the community structure of arbuscular mycorrhizal fungi was strongly affected both by soil salinity and pH [
41]. In a coastal dune ecosystem, diversity in mycorrhizal fungi was clearly segregated between the seaward (wind-disturbed) and landward (stabilized) slopes of dunes [
42]. Therefore, it was concluded that zonal distribution of both abiotic and biotic (including host plant species) factors are determinants of the fungal community structure.
In addition, apart from genetical diversity, functional variability of mycorrhizal fungi seems to be important for the outcome of the effectiveness of the symbiotic relationship. Thus, it was shown that even communities of arbuscular mycorrhizal fungi with relatively low diversity may have significant functional heterogeneity [
43]. Such characteristic features refer both to the nature of hyphal growth pattern and the intensity of mineral uptake, and they could also be important in maintaining the diversity in coastal grasslands, especially in mineral-poor soils, such as dune-associated grasslands.
3. Mycorrhizal Symbiosis in Resource Acquisition
It is generally accepted that typical mycorrhizal plants provide fungal partners with sugars and vitamins, receiving in return water and minerals, mostly N, P, and K [
44,
45]. However, terrestrial orchid species have so-called mixotrophic type of nutrition and are dependent on the mycorrhizal partner at certain stages of development and receiving sugars and vitamins from it. Nutrition of mycoheterotophic achlorophyllous plants occurs in the same way. Due to the potentially beneficial effect of the interaction on both partners, mycorrhizal symbiosis is designated as mutualistic and therefore essentially positive.
However, it must be remembered that mycorrhizal symbiosis is not always entirely mutualistic. While usually both partners benefit from the interaction, a continuum of mutualism–parasitism exists in nature [
46]. The relationship can be shifted towards parasitism due to the genetic specificity of the particular plant–fungus interaction, or it can be induced by plant developmental stage or environmental factors. Some studies performed with grassland species in controlled conditions indicate that the nature of the mycorrhizal interaction may change differentially depending on the specific situation. Typical grassland species differentially responded to mycorrhizal colonization depending on their relative abundance: dominant species
Taraxacum officinale and
Agrostis capillaris were more negatively affected by parasitic-oriented strain of
Glomus intraradices, but less abundant (subordinate) species
Prunella vulgaris and
Achillea millefolium were not negatively affected by the fungus [
47]. Thus, plant hierarchy in grasslands can be significantly affected by the presence of particular taxa of mycorrhizal fungi. Moreover, mycorrhizal association of
Hieracium pilosella was clearly beneficial, while it was parasitic for
Corynephorus canescens, especially under species competition [
48]. Both high available P concentration in soil and shade shifted mycorrhizal interaction from mutualistic to parasitic, showing that the costs of C sent to symbiont exceeded the benefits from increased mineral nutrient availability in these conditions. In this respect, it would be important to determine how the saltwater inundation characteristic of wet coastal grasslands affects the mycorrhizal dependence of different plant species relative to the mutualism–parasitism continuum, especially given the differences in salinity tolerance of various plant species.
4. Common Mycorrhizal Networks
The concept of common mycorrhizal networks (CMNs) has gained much scientific interest within recent decades [
44,
45,
49,
50,
51,
52,
53]. However, due to obvious technical difficulties, experimental evidence for existence of CMNs in nature is still scant. Instead, studies of varying degrees of complexity are conducted in different model systems under controlled conditions.
Historically, insights into mycorrhizal hyphal associations between multiple plants began with observations of the specific type of nutrition of achlorophyllous parasitic plants, mycoheterotrophy, where reduced carbon substances are received from a mycorrhizal partner, associated with an autotrophic plant [
54]. Later, with the discovery of the presence of long-branched extraradical hyphae present in both ectomycorrhizal and arbuscular mycorrhizal associations, the understanding of the potential globality and functional importance of the mycorrhizal network expanded significantly.
The importance of non-mycorrhizal plants (non-host plants) in CMNs has been recently reviewed [
55]. In particular, it was concluded that only fungal hyphae from already established mycorrhizal symbiosis can penetrate roots of non-host plants, without formation of any characteristic symbiotic structures. Usually, non-host plants are negatively affected by this type of interaction, and effects on systemic resistance are highly likely.
In contrast to unidirectional movement of mineral nutrients towards host plants, water transport in CMNs is bidirectional and changes during the day [
45]. Similarly, it is proposed that movement in signaling substances by means of CMNs can occur in different directions [
45]. Carbon cycling has been shown to occur in ectomycorrhizal CMNs but is still controversial in respect to AM networks [
50,
56]. Therefore, it seems that, apart from some specific situations (as in the case of mycoheterotophic and mixotrophic associations), CMNs are less important as a mechanism for sharing mineral resources between symbiotic plants but rather act as means for information exchange between plants. Thus far, most evidence on the importance of mycorrhizal networks in plant communication is associated with studies on ectomycorrhiza-dominated ecosystems [
51]. It has been proposed that stress-associated signals are transmitted more quickly through CMNs if compared to transfer of resources [
56]. However, the chemical nature of the signals is far from clear. Initially, it was proposed that plant hormones salicylic acid and jasmonic acid are involved in the transfer of information through CMNs [
57]. Recently, scientific information has begun to accumulate that small RNAs can be involved in important aspects of mycorrhizal symbiosis [
58,
59,
60], but evidence for their role in signaling through CMNs is still lacking.
Moreover, recently, a term “hyposphere” was coined to describe a zone of soil around mycorrhizal hyphae where release of hyphal exudates results in establishment of specific abiotic and biotic conditions [
61], forming similar differences from the bulk soil as in the case with the rhizosphere. Similar to exudates from plant roots, exudates from fungal hyphae also have an impact on bacterial diversity and abundance [
61].
Continuum of specificity of mycorrhizal fungi is an important aspect to consider in respect to development and function of CMNs [
52]. Overlap of host plant compatibility for a particular fungal strain is a critical characteristic for formation of functional CMNs, and, usually, this feature is found for dominant fungal taxa. Therefore, it is logical to assume that dynamics of plant communities are strongly dependent on functioning of CMNs, but empirical evidence for coastal grasslands is still not available. Potential mycorrhiza-related plant interactions in coastal grasslands are shown in
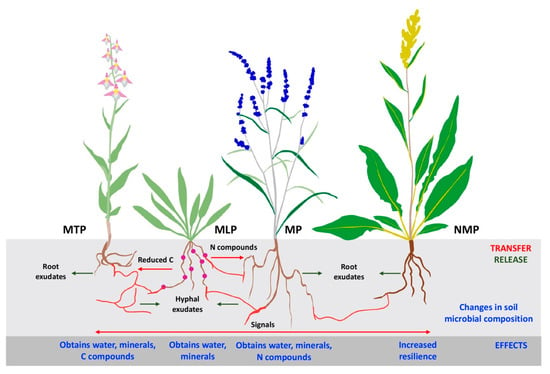
Figure 1.
Figure 1. Mycorrhiza-related interactions in coastal grasslands. MTP, mixotrophic plant species; MLP, mycorrhizal legume plant species; MP, mycorrhizal plant species; NMP, non-mycorrhizal plant species. Mycorrhizal root fragments and mycorrhizal hyphae are shown in red.
5. Mycorrhizal Symbiosis in Environmental Resilience
The issue of plant adaptation to heterogeneous environmental conditions is particularly important in grassland systems that are subject to sharp fluctuations in environmental conditions, such as coastal grasslands. The presence of mycorrhizal symbiosis in coastal plants has led to the idea of the importance of this type of symbiosis in plant adaptation. However, there has been a scientific debate on which partner of mycorrhizal symbiosis is more vulnerable to environmental constraints, plant host or fungal symbiont? Because there is a concept that mycorrhizal symbiosis is important for the plant to overcome environmental extremes, one might think that the fungal partner is the stronger side in this respect. There is no doubt that, similar to plant species specificity in tolerance to particular environmental factors, mycorrhizal fungal species and strains also differ in their ability to tolerate unfavorable conditions. Indeed, different mycorrhizal fungal species and strains show a wide range of tolerance to one of the dominant environmental constraints in coastal grasslands, soil salinity [
62]. More specifically, spore germination was delayed in the presence of NaCl, and, in some cases, the spores did not germinate at all in saline conditions, but, in others, they reached a maximum germination in the presence of 300 mM salt. Similarly, the rate of hyphal extension of some fungal taxa was even stimulated in the presence of 150 mM NaCl, but, in general, salinity inhibited hyphal growth to a varying extent.
Field studies of mycorrhizal symbiosis associated with halophytic plant species in highly saline habitats and often extremely variable moisture levels have usually revealed the presence of functionally active mycorrhizal structures in their roots, however with variable colonization intensity. Examples of such studies are from Tabriz Plain in Iran [
63], Central European salt marshes [
34] (Hildebrandt et al. 2001), Sečovlje solar salterns in Slovenia [
64], and saline soils in Hungary [
65]. Seasonal trends in mycorrhizal colonization have been assessed for halophytes in Hungarian steppe habitats [
66] and in saltwater-affected wet grassland [
11]. The results of these studies suggest that intensity of root colonization is indeed negatively affected by increasing soil salinity, but number of fungal spores does not depend on the level of salinity. It also seems that environmental factors that have an impact on host plant physiology also affect fungal symbionts.
Both diversity and composition of mycorrhizal fungi only marginally differed between halophyte and non-halophyte species growing on salt-affected coastal plains [
41]. In a typical halophyte species well-adapted to intermediate and high salinity, such as
Tripolium pannonicum (syn.
Aster tripolium), early stages of symbiotic interaction were more negatively affected by salinity in comparison to expansion of root colonization [
67]. On the other hand, seasonal changes in intensity of mycorrhizal colonization in roots of several halophyte species growing in salt-affected grassland (
Aster tripolium,
Glaux maritima,
Plantago maritima,
Trifolium fragiferum,
Triglochin maritima) showed negative dependence on fluctuations of soil salinity [
11].
A number of entirely practically oriented studies on improvement in salinity tolerance in glycophytic crop species as a result of mycorrhizal fungi application are available, indicating that the use of salt-tolerant fungal strains has great practical potential. Such strains could be isolated from the rhizosphere of salt-affected coastal grassland soils. In particular, mycorrhizal inoculation increased plant growth under saline conditions for
Gossypium arboreum [
68],
Pennisetum glaucum [
69],
Zea mays [
70],
Triticum aestivum [
71],
Lactuca sativa [
72],
Lens culinaris [
73], and
Ocimum basilicum [
74]. In some studies, it has been stressed that the fungal strains used were isolated from saline habitats [
70]. In one case, effect of two isolates of
Glomus mosseae, either form non-saline or saline soil, were compared, and it was shown that, contrary to what was initially expected, the isolate from non-saline soil had a higher capacity to alleviate negative effects of salinity [
68].
Similar experiments in controlled conditions have been performed also with some halophyte species. Thus, it was shown that mycorrhizal plants of grass species
Puccinellia distans had better growth potential under saline conditions due to enhanced photosynthesis and improved ion homeostasis [
75]. In addition, mycorrhizal symbiosis affected accumulation of osmotically active mineral elements, allowing to avoid uptake of Na [
76]. In particular, for
Trifolium alexandrinum, increased phosphorus uptake in mycorrhizal plants was associated with their better growth in saline conditions [
77]. Nine psammophilic species native to coastal sand dunes and evidently adapted only to salt spray were tested for their ability to recover after repeated seawater treatment in controlled conditions when inoculated with mycorrhizal fungus
Glomus intraradices [
78]. The intensity of the survival-promoting effect of mycorrhizal colonization was a distinctly species-specific feature. On the positive side, mycorrhizal
Ammophila arenaria plants showed less than 20% mortality after fourth application of 100% seawater concentration, while all non-mycorrhizal plants died after the third application of 100% seawater. From the worst side, mycorrhization of
Dorycinum pentaphyllum plants only marginally improved their survival under diluted seawater treatment. However, both species showed similarly high mycorrhizal dependency and high intensity of root colonization by mycorrhizal fungus.
Soil flooding, either with fresh water or seawater, represents another common environmental factor in low-lying coastal grasslands. A number of practically oriented studies on flooding tolerance of mycorrhizal plants have been performed, including seedlings of
Citrus sinensis [
79],
Prunus persica [
80,
81],
Pterocarpus officinalis [
82], and showing the overall beneficial effect of mycorrhization on flooding tolerance. Improved mineral nutrition, proline production, and suppression of ethanol production in roots during anoxic conditions were among the mechanisms responsible for growth improvement due to mycorrhizal symbiosis in flooded conditions. However, in a study with
Oryza sativa, it was shown that, while symbiosis activated the phosphorus uptake pathway in a fungal partner, it suppressed phosphorus uptake of the host plant [
83]. As a result, mycorrhizal colonization decreased shoot phosphorus content in flooded conditions, and the effect of symbiosis was negative.
Closer to the topic of this review, two grass species (
Panicum hemitomon and
Leersia hexandra) native to nutrient-poor depressional wetlands in the southeastern USA coastal plain were used in a wetland mesocosm experiment to determine if controlled water regimes affect mycorrhizal colonization as well as if colonization affects plant growth [
84]. It appears that intensity of mycorrhizal colonization decreases with increasing water levels even for species well adapted even to semi-aquatic conditions. However, mycorrhizal viability was not negatively affected, and symbiotic plants had higher phosphorus uptake even under flooded soil conditions in comparison to non-mycorrhizal plants. Mycorrhizal colonization of the same two grass species was assessed in field conditions along a hydrological gradient, and it was evident that the degree of root colonization decreased with water depth, but this did not affect number of mycorrhizal propagules in soil [
85]. However, even plants growing in permanently flooded soil retained active mycorrhizal symbiosis in roots. Similarly, the number of hyphae and spores in soil with
Zea mays plants was not affected by extended flooding in controlled conditions [
86]. Other studies also supported the idea that flooding negatively influences root colonization with arbuscular mycorrhiza, but basic symbiotic functionality is not affected [
67,
87]. Similarly, in halophyte
Aster tripolium plants, better tolerance of mycorrhizal plants to flooding was associated with improved osmotic balance and nitrogen uptake [
88].
A field study along a tidal gradient in a mangrove swamp indicated that duration of flooding period mainly affected the community structure of arbuscular mycorrhizal fungi and resulted in increased intensity of mycorrhizal colonization [
89]. When aquatic species
Polygonum hydropiper and semiaquatic species
Panicum repens grown under different hydrological regimes were compared in respect to mycorrhizal colonization and mycorrhizal community structure in natural conditions, it appeared that high flooding intensity led to a decrease in both mycorrhizal intensity and diversity level in both species [
90]. However, moderate flooding resulted in an increase in mycorrhizal colonization and fungal species richness only in aquatic species
Polygonum hydropiper. Recently, the role of arbuscular mycorrhizal symbiosis in wetland plants has been reviewed, and it was concluded that survival and development of these plants in native conditions is highly dependent on mycorrhizal colonization [
91].
It is difficult to generalize a potential role of mycorrhiza in respect to plant adaptation to soil moisture regime in coastal grasslands due to extreme variability in this factor across different coastal grassland habitats. However, it is evident that soil moisture regime is a significant determinant of both mycorrhizal community structure as well as intensity of symbiosis and its functional properties [
92]. On the other hand, there is no doubt that mycorrhizal symbiosis modulates morphological and biochemical adaptations of drought-stressed plants, as summarized in the recent reviews [
93,
94]. As mycorrhizal colonization usually results in formation of induced systemic resistance of host plants [
95], further studies of specific responses to drought in mycorrhizal vs. non-mycorrhizal coastal grassland plants are needed.
This entry is adapted from the peer-reviewed paper 10.3390/land12061209