1. Capacity of Photosynthesis and Photosynthesis-Related Processes across Growth Light Intensity in Floating Aquatic Plants versus Land Plants
1.1. Photosynthetic Capacity
Acclimatory upregulation of light- and CO
2-saturated photosynthetic capacity, with a much greater capacity on a leaf area basis, is seen in high- compared to low-light-grown plants of terrestrial species (
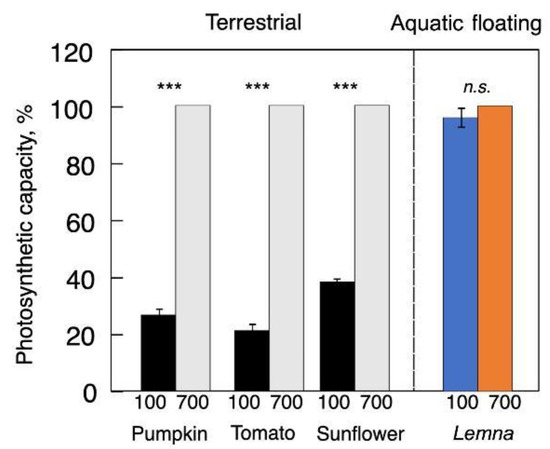
Figure 1; [
17]). In contrast, floating aquatic plants like those in the genus
Lemna exhibit constitutively high levels of the key photosynthetic protein Rubisco [
46] and photosynthetic capacity across a wide range of growth light intensities (
Figure 1; [
41,
47]).
Figure 1 shows that
L. gibba exhibited up to 50% protein of dry biomass [
40] present in the as Rubisco [
46], and an associated capacity of photosynthetic oxygen evolution even in plants grown in low light [
40,
48]. Martindale and Bowes [
49] showed that, whereas only a small fraction of Rubisco is activated under low light, all of this Rubisco can be rapidly activated upon exposure to high light (and without need to produce additional Rubisco-rich cell layers) to support a high photosynthetic capacity in low-light-grown duckweed.
Figure 1 shows such high photosynthetic capacities of leaves of land plants and fronds of duckweed–each developed under either low or high growth light intensity. In other words, duckweeds maintain what can be considered sun-plant characteristics with respect to photosynthetic capacity even when grown under low light intensity.
The low maximal photosynthetic capacity of leaves of land plants grown under low light intensity is consistent with a much lower Rubisco level in these leaves compared to leaves grown under high light. It should be noted that the extent of photosynthetic acclimation of terrestrial plants to changes in the light environment varied with species-dependent foliar anatomical constraints [
50,
51] and with whether transfer from low to high light occurred before or after a leaf was fully expanded [
52].
Figure 1. Light- and CO
2-saturated rate of photosynthetic oxygen evolution (on a leaf or frond area basis) for pumpkin, tomato, sunflower (left side), and
Lemna gibba (in color, right side) grown under 100 and 700 µmol photons m
−2 s
−1. Values are given as percentages of maximum values in plants grown under 700 µmol photons m
−2 s
−1 for each species (pumpkin = 50, tomato = 53, sunflower = 59, and
L. gibba = 33 µmol O
2 m
−2 s
−1). Significant differences are signified by asterisks (***,
p < 0.001).
n.s. = not significant. For original data and further statistical information, see Stewart et al. [
48] and Polutchko et al. [
53].
1.2. Vascular Infrastructure for Sugar and Water Transport in Land Plants
Acclimatory upregulation of photosynthetic capacity (as the capacity of sugar production) in land plants is accompanied by upregulation of the vascular infrastructure for foliar sugar export and water import in response to higher growth light intensity (
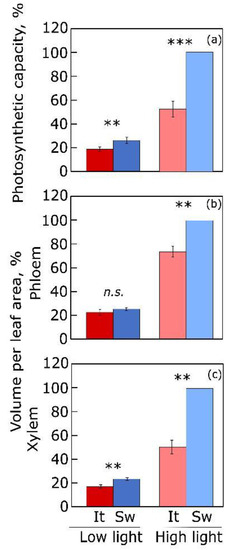
Figure 2; [
54,
55,
56];
Section 5 below addresses a similar concomitant upregulation of maximal photosynthetic capacity and foliar vascular infrastructure in response to cool temperature). Furthermore, such acclimatory responses of the individual leaf/plant vary in degree because of genetic adaptation to different habitats of origin for a species or ecotype–with different light supply, temperature, and water availability [
4,
54].
Figure 2 shows substantial concomitant acclimation of maximal photosynthetic capacity to growth light environment in
Arabidopsis thaliana ecotypes from Sweden and Italy as well as the greater degree of this response in the Swedish ecotype [
54,
56].
Figure 2. (
a) Photosynthetic capacity (light- and CO
2-saturated rate of photosynthetic oxygen evolution) and volume per leaf area (as the mathematical product of cross-sectional area per vein × vein length per leaf area) for (
b) phloem and (
c) xylem cells in minor veins from leaves of two ecotypes (from Sweden, Sw; from Italy, It) of
Arabidopsis thaliana grown under low (100 µmol photons m
−2 s
−1) and high (1000 µmol photons m
−2 s
−1) light (9-hour photoperiod) under controlled conditions (20 °C leaf temperature). Data are expressed as percentages of maximum values for the Swedish ecotype grown under high light (photosynthetic capacity, 71.9 µmol O
2 m
−2 s
−1 and 4057 and 1390 mm
3 m
−2 for volume per leaf area of phloem and xylem, respectively). Significant differences are signified by asterisks (**,
p<0.01; ***,
p<0.001).
n.s. = not significant. For original data and further statistical information, see Stewart et al. [
56].
Because of their diminutive size, their intimate contact with water, and the fact that they consist mainly, or exclusively (in the rootless
Wolffia arrhiza), of photosynthetic tissue [
28], Lemnaceae presumably require minimal vascular tissue for water and sugar transport [
57]. Nutrients are taken up through the lower epidermis in contact with water under ample nutrient supply (with increasing contributions from rootlets extending into the water under low nutrient supply; [
58]). Vascular tissue is absent in the genera
Wolffia and
Wolffiella and exhibits a low degree of complexity in
Spirodela, Landoltia, and
Lemna [
28,
59], with plasmodesmatal connections between adjacent cells [
60] fulfilling the role of transporting water and carbohydrate. Overall, the morphology and anatomy of duckweeds is thus thought to be well adapted for these small plants’ habit of floating on water surfaces and showing a potential for rapid growth during relatively short growing seasons [
28,
29]. Kim (2007) [
60] specifically highlights the contributions of a plant body packed with chloroplasts throughout and with abundant plasmodesmata, air channels, as well as mechanisms that support expedient separation of daughter fronds from mother fronds during rapid vegetative propagation.
1.3. Antioxidants in Land Plants
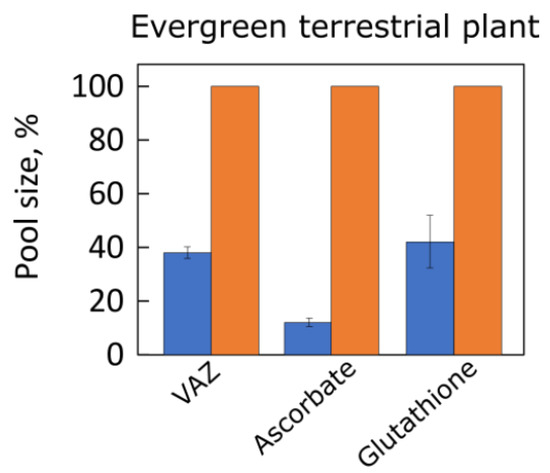
Figure 3 shows pronounced acclimatory differences in the pool size of the photoprotective xanthophyll cycle pigments (violaxanthin, antheraxanthin, and zeaxanthin = VAZ) as well as the antioxidant metabolites ascorbate and glutathione in response to growth of a terrestrial plant under high versus low light [
61]. Zeaxanthin protects both plants and humans against oxidative damage
. Ascorbate (the human vitamin C) supports VAZ pool conversion to zeaxanthin [62] and also participates in the detoxification of hydrogen peroxide to water by ascorbate peroxidase in the chloroplast (Mehler reaction; [63]). Glutathione supports reduction (recycling
) of vital thiol-containing antioxidant enzymes in the chloroplast and other cellular compartments [64]. Additional components of antioxidant systems exhibited similarly pronounced acclimation to growth light intensity [22,61].
Figure 3. Pools of the xanthophyll cycle pigments (VAZ), ascorbate, and glutathione in leaves of the evergreen groundcover
Vinca major grown under 100 (blue bars) and 1200 (orange bars) μmol photons m
−2 s
−1. Data are given as percentages of maximum values in plants grown under 1200 μmol photons m
−2 s
−1 for each compound (VAZ = 91 mmol mol Chl
−1, ascorbate = 21 µmol g
−1 fresh weight, and glutathione = 968 nmol g
−1 fresh weight). All differences between leaves grown under low versus high light were statistically significant. For original data and statistical information, see Grace and Logan [
61]. A, antheraxanthin; V, violaxanthin; Z, zeaxanthin.
Whereas the level of antioxidant metabolites and other antioxidant systems still needs to be characterized across a range of growth light intensities in Lemnaceae, available evidence indicates high levels of carotenoids (except for zeaxanthin), antioxidant vitamins, and mineral cofactors of antioxidant enzymes [
65] as well as multiple phenolics with antioxidant/anti-inflammatory properties [
66,
67,
68,
69] even in plants grown under very low light intensity [
70]. Similarly, [
40,
47,
48] reported high levels of the carotenoids
β-carotene (provitamin A) and lutein as well as vitamin E in duckweed plants grown under low light intensity.
2. Comparison of How Quickly Substantial Zeaxanthin Is Formed upon Sudden Transfer from Low to High Light in Land Plants versus Floating Aquatic Plants
2.1. Changes in Xanthophyll Cycle Pool Size and Conversion in Terrestrial Plants upon Transfer
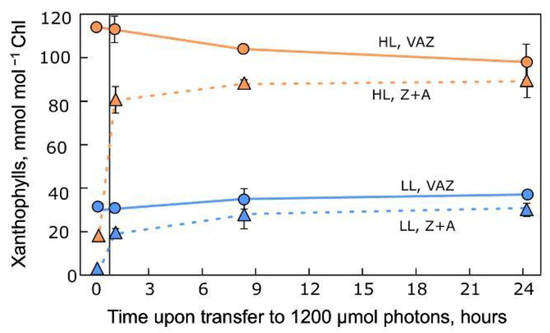
The example depicted in Figure 4 illustrates that, compared to high-light-grown individuals, low-light-grown individuals of the same (land plant) species were unable to quickly accumulate large amounts of the photoprotective components (zeaxanthin and antheraxanthin, Z+A) of the xanthophyll cycle upon exposure to high light for 24 hours following 12 hours of darkness. The VAZ pool was several-fold lower in individuals of this same plant species when grown under low (versus high) light and remained at these different levels through the high-light treatment.
Figure 4. Changes in xanthophyll cycle (VAZ) pool and Z+A levels in leaves of evergreen
Schefflera arboricola grown under low (LL) or high (HL) light intensity (100 or 1200 µmol photons m
−2 s
−1, respectively) and then subjected to 24 hours of exposure to 1200 µmol photons m
−2 s
−1. Data from Demmig-Adams et al. [
71]. A, antheraxanthin; V, violaxanthin; Z, zeaxanthin.
Similar results were reported for a range of additional terrestrial species, including both annuals and evergreens [
72,
73]. For example, plants of several terrestrial species grown in non-excessive light and suddenly transferred to 1100 µmol photons m
−2 s
−1 increased their VAZ pools only gradually over the course of a whole week [
72]. In the case of cotton (as an example for a fast-growing annual species), new leaves developed over the course of a week and exhibited features of high-light-grown leaves [
72]. Evergreen species did not develop new leaves over this period. In contrast, Lemnaceae can form a whole new plant (by vegetative clonal formation of daughter fronds from a mother frond) in just a day or two. This example further illustrates that, unlike natural environments with light fluctuations, a growth environment with constant low light intensity fails to prepare terrestrial plants for coping with a sudden change in growth light intensity (see also [
10,
18,
22]).
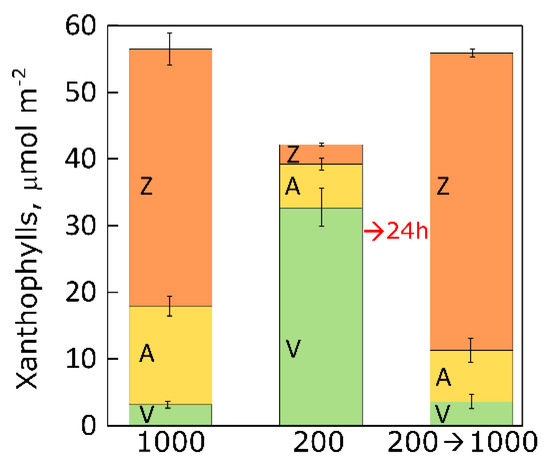
2.2. Changes in Xanthophyll Cycle Pool and Conversion in Lemna upon Transfer
The example depicted in
Figure 5 shows that
L. minor grown in low light (similar light conditions to those of the above-mentioned land plants) (i) had VAZ pools that were not much lower (about 25%) than those of plants grown in high light, (ii) were able to increase their VAZ pool to the size of that in plants grown at high light within 24 hours (rather than over several days) upon sudden transfer from low to high light, and (iii) accumulated as much zeaxanthin by 24 hours post-transfer as plants that had developed in continuous high light. This finding demonstrates that
L. minor did not require extended acclimation after a sudden increase in light intensity (see above). Moreover, there were also no signs of heat damage in
L. minor upon sudden transfer from low to high light, which could be due to duckweed fronds’ high surface-to-volume ratio and contact with thermostable water (see above). In contrast, land plants grown in low light typically possess insufficient vascular infrastructure (see
Figure 2 above) and leaf pore (stomatal) density [
74] to provide the necessary evaporative cooling upon sudden transfer from low to high light. In other words, this example suggests that floating aquatic plants–perhaps due to genetic adaptation to fluctuating environments–can cope with a sudden dramatic change in growth light intensity without requiring prior exposure to at least some periods of increased light intensity during individual plant development.
Figure 5. Xanthophyll cycle components in L. minor grown under three light (16 h light/8 h dark) conditions, (i) high light (1000 µmol photons m−2 s−1) for four days, (ii) low light (200 µmol photons m−2 s−1) for four days, and (iii) 200 μmol photons m−2 s−1 for three days followed by transfer (→) to continuous high light (1000 μmol photons m−2 s−1) for 24 hours. V = Violaxanthin, A = antheraxanthin, and Z = zeaxanthin.
3. Acclimation to Other Environmental Factors in Land Plants and Lemnaceae
3.1. Temperature Acclimation in Land Plants
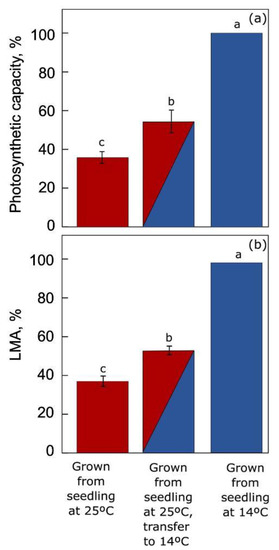
Figure 6 shows (i) the pronounced extent of acclimation to warm or cool growth temperature in
A. thaliana plants grown under these contrasting conditions from seedling stage as well as (ii) the limited extent of acclimation reached by warm-grown plants suddenly transferred to lower temperature [
3]. This terrestrial winter annual (but not summer annuals; [
75]) thus exhibited pronounced acclimatory adjustments in leaf form and function, including a much greater maximal photosynthetic capacity and leaf mass per area (as well as greater foliar sugar-export infrastructure, [
4,
54,
75,
76]) in leaves that developed under cool versus warm temperature. Transfer of mature warm-grown leaves to cool temperature for one week did produce some acclimatory adjustment in these features but was unable to lead to the full acclimation seen in leaves that developed under cool temperature. This result further demonstrates the need for multi-day acclimation to the environment in terrestrial plants and the partial, but limited, ability of their mature leaves to reach full acclimation of form and function upon sudden transfer.
Figure 6. Comparison of (
a) light- and CO
2-saturated photosynthetic oxygen evolution and (
b) leaf dry mass per area (LMA) for plants of an
A. thaliana Swedish ecotype grown either at warm (25 °C; red) or cool (14 °C; blue) leaf temperature from seedling stage or suddenly transferred to 14 °C for one week (red/blue) after growth (with development of fully mature leaves for characterization) at 25 °C. Data are given as percentages of the maximum values for
A. thaliana grown from seedling stage under 14 °C (75 µmol O
2 m
−2 s
−1 and 62 g m
−2, respectively, for photosynthetic capacity and LMA). Significant differences among the three treatments are signified by lower-case letters. For original data and further statistical information, see Adams et al. [
3].
3.2. Response to Low or High Nitrogen Supply in Lemnaceae
As shown above, development of a new leaf in an altered environment is often needed for full acclimation of form and function. In terrestrial species, new leaves require 5 to 7 days to mature in fast-growing annuals and longer in woody and evergreen species. In contrast, Lemnaceae can form whole new plants (daughter clones from mother clones) in 1 to 2 days (for morphological information, see [
29]). This difference would be expected to reduce the time required for acclimation to a new environment. More research is needed to compare the need for acclimation to environmental factors other than light between Lemnaceae and terrestrial plants.
The response of Lemnaceae to the level of nitrogen supply is defined by their ability to (i) acquire nitrogen from endophytic bacterial symbionts (e.g.,
Rhizobium lemnae) and other N
2-fixing microorganisms [
77] and (ii) retain large amounts of vegetative storage protein as an endogenous source of nitrogen for new growth. For example,
L. minor persisted for several months over the summer with green fronds [
40,
47] in a pond with negligible levels of nitrogen in the water (as either nitrate or ammonia; Christine M. Escobar, personal communication). These plants exhibited a photosynthetic capacity of 15 ± 2 µmol O
2 m
−2 s
−1 (Jared J. Stewart, unpublished data). This effect is likely related to the ability of duckweeds to exhibit some growth even under negligible nitrogen supply in the medium by utilizing nitrogen provided via bacterial N
2 fixation [
77]. Similarly,
L. minor exhibited a remarkably high photosynthetic capacity (of 66.6 ± 2.3 µmol O
2 m
−2 s
−1) after 4 days upon transfer from a medium with ample nutrient supply (1/2 strength Schenk and Hildebrandt medium; bioWORLD, Dublin, OH, United States; [
78]) to a medium with low nutrient supply (1/20 strength Schenk and Hildebrandt medium).
Whereas the above features suggest an ability of Lemnaceae to buffer against low nutrient supply in the medium, there is also evidence for photosynthetic acclimation in response to increased nutrient supply in
Lemna on a time scale similar to what is seen in land plants for acclimation to light environment. Photosynthetic capacity of fronds collected from the above-described pond with negligible nitrogen content increased to an astounding rate (on a frond area basis) of 100 µmol O
2 m
−2 s
−1 over 14 days upon transfer to ample nutrient supply (1/2 strength Schenk & Hildebrandt medium). This increase in photosynthetic capacity on a frond area basis was associated with a concomitant matching increase in dry biomass per frond area, such that photosynthetic capacity on a dry biomass basis remained the same. This response is similar in its time course and features to that of land plants for a comparison of low- versus high-light-grown plants, where photosynthetic capacity and dry biomass per leaf area are greater in high versus low light in proportion with each other, resulting in an unchanged photosynthetic capacity on a dry biomass basis [
79]. It can be concluded that
L. minor did undergo acclimatory changes in photosynthetic capacity in response to a sudden increase in nutrient supply in the medium. This conclusion is also consistent with the finding of acclimatory adjustment in root length in response to low nutrient supply [
58] as well as acclimatory adjustments in response to different levels of copper exposure [
80] in
L. minor.
3.3. Maintenance of Light-Harvesting Capacity in Lemnaceae despite Growth Arrest
Lemnaceae exhibit a notable ability to maintain a high light-harvesting capacity (green fronds) even with negligible nitrogen supply from the medium (in an outside pond), which differs from the response of annual land plants like spinach but is similar to the response of evergreen land plants. Under low (but not negligible) nutrient supply, all but the youngest leaves of spinach turned yellow [
1] with apparent transfer of nitrogen from older to younger leaves. This yellowing was seen when plants were watered daily with a (low-nitrogen) medium containing 0.25 mM rather than (a nitrogen-replete) medium with 14 mM nitrate [
1]. In contrast, soybean (as a terrestrial species with N
2-fixing symbionts as seen in
Lemna) exhibited some growth even in the absence of nitrogen supply in the soil [
81].
Lemna minor did appear to arrest clonal divisions in the nutrient-depleted pond once the pond was completely covered with
Lemna. Such an effect of crowding when coverage is high has been documented under controlled conditions as well. Duckweed growth rates are highest when density of coverage is low and drop off when high percentages of coverage are reached (see [
82]). It appears that in clonal floating aquatic plants, growth arrest is related to competition for nutrients and space rather than for other abiotic factors (e.g., light).
Overall, these responses to variable nutrient supply and temperature in Lemnaceae represent a combination of the contrasting responses of annual and evergreen land plants in combining the potential of terrestrial annuals for very fast growth under favorable conditions with the ability of terrestrial evergreens to stay green while exhibiting growth arrest under unfavorable conditions. These findings further support the notion that floating aquatic plants possess unique combinations of traits that may be a result of the environment in which they evolved.
This entry is adapted from the peer-reviewed paper 10.3390/plants12101928