Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Subjects:
Food Science & Technology
|
Chemistry, Medicinal
I3C is an important phytochemical contained in cruciferous vegetables and is able to exert various activities among which are cardioprotective, antioxidant, anti-inflammatory, antiangiogenesis, and antimicrobial activities, the promotion of tumor cell apoptosis and, more recently, an important inhibition of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS)-CoV-2 viral egression, including the Omicron variant.
- Indole-3-carbinol
- phytochemical
- natural products
1. I3C Biosynthesis
With regard to the environment of plants, the role of glucosinolates is to protect them. Upon damage to plant tissues, they undergo enzymatic hydrolysis and are transformed into bioactive molecules toxic to herbivores and pathogens. When plant tissues are damaged, the β-thioglucosidase [17] enzyme myrosinase [18] can hydrolyze the thioglucosidic bond in the structure of the glucosinolates [2]. In particular, I3C is obtained from the degradation of the glucosinate glucobrassicin. This process is carried out through the formation of unstable intermediates thiohydroximate-O-sulfonate and 3-indolylmethyl isothiocyanate, which is finally converted into the desired compound I3C (Figure 3).
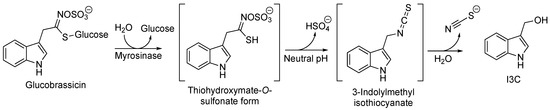
Figure 3. Biosynthesis of I3C starting from glucobrassicin.
2. I3C Synthetic Methodologies
Among the numerous synthetic methodologies followed to obtain I3C, those with the best characteristics to be reproduced on a very large scale (the first illustrated) and the one with very high yield were selected.
A classical method to obtain I3C requires highly basic conditions such as the reduction of the corresponding aldehyde with sodium boron hydride (NaBH4) [19,20], as follows in Scheme 1.
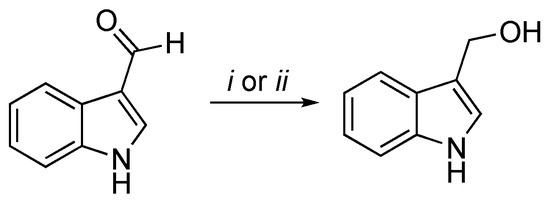
Scheme 1. Reagents and conditions: (i) NaBH4, MeOH, 0 °C, 30′; (ii) NaBH4, EtOH/THF, rt, 3 h.
The second scheme describes the method to obtain I3C from indole proposed by Downey et al. [21] (Scheme 2). Indoles undergo Friedel–Crafts addition to aldehydes in the presence of trimethylsilyl trifluoromethanesulfonate (TMSOTf) and a trialkylamine to form 3-(1-syloxyalkyl)indoles. The reaction is quenched by the addition of pyridine followed by deprotection under basic conditions with tetrabutylammonium fluoride (TBAF) and provides the free desired product. This method prevents the formation of bisindolyl methanes, a thermodynamically favored process typically observed when indoles react with aldehydes under acidic conditions.
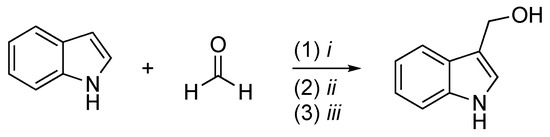
Scheme 2. Reagents and conditions: (i) TMSOTf, i-Pr2NEt, Et2O, N2, −78 °C, 1 h; (ii) pyridine; (iii) TBAF, THF, rt, 5′.
As mentioned before, I3C is a well-researched and interesting compound due to its extensive biological properties and may activate multiple antiproliferative cascades [22].
For an extended period, I3C has been widely explored for potential roles in different cellular mechanisms, including the suppression of cell cycle progression, blocking of cancer cell migration, the promotion of apoptosis, and the inhibition of tumor growth [23,24,25]. Although it interacts with different pathways, it has been proposed that I3C and its synthetic derivatives may influence human cells by directly inhibiting them with specific enzymatic target proteins. Specifically, they are potent natural inhibitors of homologous to E6AP carboxyl-terminus (HECT) family members of E3 Ubiquitin-Ligase, emphasizing the potential importance of I3C in developing highly potent and stable anti-cancer molecules, and not only [26]. I3C and its derivatives are also able to inhibit neuronal precursor cell-expressed developmentally downregulated 4 (NEDD4) and WW domain-containing ubiquitin E3 ligase 1 (WWP1) ubiquitination enzymatic activity through the interaction with their catalytic subunit [26,27,28,29].
Since I3C affects many cellular mechanisms, it is not surprising to find proposed applications in the treatment or prevention of different cancer types (e.g., breast, ovarian, prostate, lung, liver, and colon) and in other diseases [30,31,32,33,34,35,36,37,38]. To date, it has been tested in many clinical trials for the prevention and treatment of obesity, chronic inflammation, lupus erythematosus (SLE), and breast, colon, and prostate cancer [39]. Details about all of the applications of the potential of I3C and its derivatives in different pathologies are reported below.
3. I3C Antitumor Activity: In Vitro, In Vivo, and Clinical Studies
Cancer is one of the leading causes of disease and death worldwide and is defined as uncontrollable and abnormal cell growth [40]. In breast cancer, gland cells grow abnormally and uncontrolled, resulting in the development of a malignant and metastatic phenotype [41]. It is divided into four molecular subtypes based on the expression of the estrogen receptor (ER), progesterone receptor (PgR), and the epidermal growth factor receptor 2 (HER2): two are characterized by the presence of hormone receptors, which provide hormonal therapeutic treatment; then, there is the HER2 subtype, which constitutes about 20% of all cases and is a biologically treatable tumor with some monoclonal antibodies specific to the substance used; finally, there is the triple-negative tumor (it has no receptors for either hormones or HER2), which is sometimes more aggressive and is subject to studies aimed at specific therapies [42].
Although conventional treatments are clinically effective against breast cancer, they present severe side effects associated with the development of resistance and high toxicity in healthy cells leading to a poor prognosis [40]. Therefore, the discovery of new treatments and in particular, the use of natural compounds is one of the primary goals in the field of breast cancer biology, due to therapeutic potential with minimum side effects compared to the traditional methods such as chemotherapy and radiotherapy [43].
In this context, different studies have been conducted on different phytochemical compounds that have brought to light significant antitumor effects, preventing malignant neoformations, inhibiting the metastatic process, and also optimizing first-line anticancer therapies [41]. For many years, several studies have highlighted the potential benefits of the phytochemical compound I3C (and its derivatives; in particular, DIM) as promising antitumoral options with negligible toxicity [23,44,45,46], due to the fact that many of these antiproliferative responses are selectively controlled by I3C-activated pathways. Now, we will illustrate in molecular detail how I3C and its derivatives exert their antitumor activity in the human cancer cell.
Regardless of the type of tumor, I3C triggers DNA repair, cell-cycle arrest, apoptosis, disruption of cell migration, and modulates hormone receptor signaling [47,48,49]. The first identified I3C target protein was serine protease elastase [50]. In human breast cancer, it has been observed that inhibition of elastase activity by I3C prevents CD40 cleavage, resulting in the disruption of the NFkB-dependent cell cycle and proliferation [51,52].
Moreover, I3C and its derivatives also play a fundamental role in DNA damage repair processes by influencing the expression of tumor suppressor genes such as BRCA1 and BRCA2 and DNA repair proteins (i.e., RAD51, which regulates responses in the case of DNA damage) [53,54]. In all estrogen-dependent tumors (including breast cancer), I3C inhibits cyclin-dependent kinase (CDK) expression, resulting in the stopping of the cell cycle in the G1 phase. In particular, I3C has also been reported to induce apoptosis in the PC-3 human prostate cancer cell line by inhibiting the activation of serine/threonine kinase Akt [55,56]. In addition, I3C has the ability to counteract the metastatic process, tumor angiogenesis, and migration of cancer cells, inhibiting CDK6 with the consequent blockage of cell growth [57]. These observations suggest that I3C may mediate its anti-proliferative effects by directly interacting with other classes of target proteins with enzymatic activities.
More recently, through the identification of genes involved in tumorigenesis and cancer susceptibility, it was found that members of the HECT family of E3 ubiquitin ligases were often over-expressed in human cancers. They display oncogenic properties through the ubiquitin-dependent regulation of several protein substrates such as phosphatase and tensin (PTEN) homolog, a negative regulator of phosphatidylinositol-3,4,5-trisphosphate and the Akt/PKB signaling pathway [29]. Specifically, two members of a subgroup of HECT-E3 ligases, known as C2-WW-HECT (NEDD4-like), have been identified to be most involved in cancer: NEDD4 and WWP1. They are characterized by WW domains that act on protein–protein interactions by recognizing Pro-rich and Ser/Thr-Pro phosphorylated patterns. These domains provide scaffolding to recruit protein substrates and regulators [26,27,28].
Quirit et al. have demonstrated through in vitro assay that I3C directly inhibits NEDD4 ubiquitination activity. Moreover, protein thermal shift assays, in combination with the in silico binding simulations and crystallographic structure of NEDD4, showed the binding involvement to the purified catalytic HECT domain of NEDD4 [26]. Likewise, due to its structural similarity to NEDD4, WWP1 was also demonstrated to be inhibited by I3C. It is interesting to note that the MST (microscale thermophoresis) binding assay found that I3C binds to the WWP1 HECT domain with a dissociation constant much higher than that related to NEDD4 [27].
In addition to directly influencing specific molecules, I3C (and DIM) also seem to be able to regulate gene expression by modulating DNA methyltransferases, histone deacetylases, miRNAs, lncRNAs, and some transcription factors such as the aryl hydrocarbon receptor (AhR), ER, and NF-kB [58,59,60]. In fact, we know, for example, that DNA methylation involves the silencing of tumor suppressor genes, an important mechanism for developing target molecules for chemoprevention, and able to modulate the methylation of these genes. In this regard, it has been seen that the inhibition of DNMT (DNA methyl transferase) by DIM has shown a lower expression of oncogenes and an increase in tumor suppressor genes [61]. This mechanism has emerged from some experiments on mouse models and on human prostate cancer cells, on healthy prostate cells (PrEC), as well as on androgen-dependent (LnCAP) or androgen-independent (PC3) prostate cancer [62].
These results suggest that I3C could provide a starting point to develop highly potent anticancer compounds based on target-protein interactions to have a more potent antiproliferative response in human breast cancer cells (and also for other types of cancer) compared to other inhibitors [52].
In recent years, the demand for natural products with prophylactic action against a series of diseases and their adverse reactions is increasing. According to some studies, I3C exerts a potential prophylactic action either directly or through its metabolites [63].
Recently, Lee et al. identified, in either Myc-driven or PTEN heterozygous mice, the reactivation of PTEN due to a pharmacological inactivation of WWP1 by I3C, leading to the potent suppression of tumorigenesis driven by the PI3K-Akt pathway [27]. Moreover, PTEN deletion by CRISPR-Cas9 in Hi-Myc tumor organoids conferred partial resistance to I3C, supporting that I3C exerts its function in a PTEN-dependent manner. In addition, an I3C pharmacokinetic analysis in male C57BL/6 mice after intraperitoneal administration at 20 mg/kg every day or every other day was carried out. These results support again that I3C targets WWP1 E3 ligase, triggering reactivation of the PTEN tumor suppressive function, and promoting its plasma membrane recruitment, and suppression of MYC-driven tumorigenesis in vivo. These findings highlight the use of I3C as a potential therapeutic strategy to reactivate PTEN (through WWP1 inhibition) and to treat all of those patients with multiple tumors or other diseases associated with PTEN germline mutations and for cancer prevention, through the targeting of WWP1-PTEN axis pharmacologically [27].
There are also several studies carried out on mouse models to test the effectiveness of I3C and its derivatives. Studies on immunodeficient mice, such as the naked mouse or the non-obese diabetic (NOD) mouse, in which human cancerous cell lines (xenotransplantations) are implanted and follow the dietary administration of I3C (or DIM), have observed the inhibition of tumor cell proliferation over time [64,65,66]. Again, following the implantation of human cancer cell lines in SCID (severe combined immunodeficiency) mice (NOD. CB17-Prkdcscid/SzJ), the efficacy of I3C in comparison to that of DIM from food in inhibiting tumor growth has been analyzed: both tumor size and doubling time of human T-ALL CCRF-CEM cell xenografts in these mice were significantly affected by 100 ppm of dietary DIM, while they were sensitive to 500 and 2000 ppm of I3C from the diet [64]. The chronic administration of I3C in rodents has led to the development of hepatocarcinogenesis [67,68], while in the infantile model, it led to a high decrease of liver tumors induced by diethylnitrosamine [69]. Some studies on colon cancer have brought to light activity of chemoprevention of I3C and DIM according to an AhR-dependent modality, also susceptible to the microbial production of indole AhR ligands starting from the metabolism of dietary tryptophan [70,71]. For the analysis of breast cancer in rats induced by DMBA (7,12-dimethylbenzanthracene) or MNU (N-methylnitrosourea), a direct-acting carcinogen, I3C, unlike DIM, has been shown to be effective in chemoprevention [72]. All of this evidence justifies the ability to attribute I3C chemopreventive activity to DIM in each study performed. Protection against TBD-dependent in female, but not male, offspring originating from mothers fed I3C during gestation required the expression of ERβ [73]. These results would be in line with the chemoprevention of I3C and ER-dependent DIM in breast cancer [23,74,75,76,77,78,79,80,81].
Preclinical studies of I3C as chemopreventive agents have yielded excellent results [24,82,83]. For example, in addition to liver, breast, and colon cancer, dietary I3C leads to a decrease in lung carcinogenesis caused by the specific nitrosamine contained in tobacco, 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone, PAH (pulmonary arterial hypertension), and BAP (broader autism phenotype) [84].
Since numerous in vitro and in vivo studies have shown its powerful chemopreventive action in different cancers, I3C has also been the subject of some promising clinical studies concerning cancer therapy. In a double-blind placebo-controlled study, the ability of I3C to prevent the onset of breast cancer was evaluated [85]. The study involved 60 women aged 22 to 74 years who received doses of 50 mg or 400 mg of I3C and in no case did adverse effects occur except in patients with a history of elevated aminotransferase levels. The study showed that I3C exerts a preventive effect on breast cancer and all forms of estrogen-dependent cancer [85]. It is important to consider that breast cancer in women has been placed in the foreground following the proven activity of I3C (and DIM) to act on CYP1-dependent estrogen metabolism. With regard to the metabolites of estradiol (E2), 2-hydroxyestrogen has a lower pharmacological activity, in contrast to 16-α-hydroxyestrogen, which maintains estrogenic activity. The ratio (2-hydroxyestrogen)/(16-α-hydroxyestrogen) to date is considered a biomarker for the identification of the risk of an estrogen-dependent tumor and for cervical intraepithelial neoplasia (CIN), such that clinical studies carried out with I3C (and DIM) have determined an increase in the ratio between the two estrogens, and therefore, the prevalence of the non-estrogenic metabolite, and, in the specific case of CIN, an improvement in the pathology.
I3C (and DIM) also affect the action of flavin-containing monooxygenases, i.e., a superfamily of enzymes contained in the endoplasmic reticulum of tissues such as the liver, lung, intestine, and kidney, which catalyze redox reactions in which NADPH is identified as a reducing agent and, at the same time, an O2 atom is added in a substrate while the other is used for the formation of H2O [86]. Humans express five FMOs (FMO 1–5) but in mammals, excluding primates, FMO1 is the main form present in the liver capable of metabolizing a wide range of xenobiotics [87]. Specifically, dietary I3C (and DIM) inhibits FMO1 only in rats and FMO3 in humans [88,89,90,91,92]. The ratio of FMO/CYP-mediated metabolism of DMA was reduced in a dose-dependent manner following the intake of dietary I3C (and DIMs). When considering (S)-nicotine, the rate of metabolism operated by CYP did not change with diet, while N-oxygenation was significantly inhibited, and no forms of N-oxide were detected in liver microsomes of rats fed 2500 ppm I3C (or 1000 ppm DIM). Tamoxifen, an estrogen receptor antagonist ER used in chemoprevention or treatment of ER-dependent breast cancer, allows limited use from ovarian toxicity due to the hydroxylation reaction at position 4 [93]. Again, the N-oxidized form is the eliminable form and the inhibition of N-oxide metabolism is likely to increase the toxicity of both (S)-nicotine and tamoxifen [94].
However, the actual clinical and preclinical I3C concentrations as well as the consumption of large quantities are still unclear; a recent paper published by Centofanti et al. evaluated potential adverse in vivo toxicity effects through two different routes of administration [intraperitoneally (i.p.) and intragastrically (i.g.)] in both male and female mice, because in females, the effect of I3C may be subject to hormonal changes. This analysis showed a different tolerability dose in males and females to be considered for any future clinical trials in which I3C will be used. Overall, below 550 mg/kg for i.g. and 250 mg/kg for i.p. values, I3C induces neither death nor abnormal toxic symptoms, as well as no histopathological lesions [11].
It is always important to remember that the doses used in preclinical and clinical studies are significantly higher than the doses that can be derived from food sources; for this reason, supplementation is essential to achieve the same health effects attributed to I3C (and DIM).
Finally, it is also important to highlight some controversies that emerged from different studies on the cellular effect of high I3C concentration. Rather than I3C, this effect should be attributed to its DIM derivative(s), because I3C rarely, if ever, is detected in the blood after oral ingestion [95]. Moreover, it has been proven that in humans, a significant portion of DIM is actually present in the form of its metabolites, having an unknown pharmacological activity [96].
Although I3C and DIM have been subjected to clinical trials in humans, essentially to investigate their efficacy against breast and prostate cancer [59,95,97,98,99,100,101,102], different epidemiological studies have been carried out on the population that have brought to light a negative relationship between the intake of cruciferous vegetables and some cancers [83,103,104,105,106].
Notably, except for one case, these clinical studies consider the therapeutic potential of DIM in cancer and not the chemopreventive activity, which in fact requires double-blind, placebo-controlled studies in disease-free subjects to confirm its potency and efficacy as a supplement.
This entry is adapted from the peer-reviewed paper 10.3390/ph16020240
This entry is offline, you can click here to edit this entry!