Diabetic foot ulcers cause great suffering and are costly for the healthcare system. Normal wound healing involves hemostasis, inflammation, proliferation, and remodeling. However, the negative factors associated with diabetes, such as bacterial biofilms, persistent inflammation, impaired angiogenesis, inhibited cell proliferation, and pathological scarring, greatly interfere with the smooth progress of the entire healing process. It is this impaired wound healing that leads to diabetic foot ulcers and even amputations. Therefore, drug screening is challenging due to the complexity of damaged healing mechanisms. The establishment of a scientific and reasonable animal experimental model contributes significantly to the in-depth research of diabetic wound pathology, prevention, diagnosis, and treatment. In addition to the low cost and transparency of the embryo (for imaging transgene applications), zebrafish have a discrete wound healing process for the separate study of each stage, resulting in their potential as the ideal model animal for diabetic wound healing in the future.
- zebrafish
- diabetic wound
- delayed wound healing
- model animal
1. Introduction
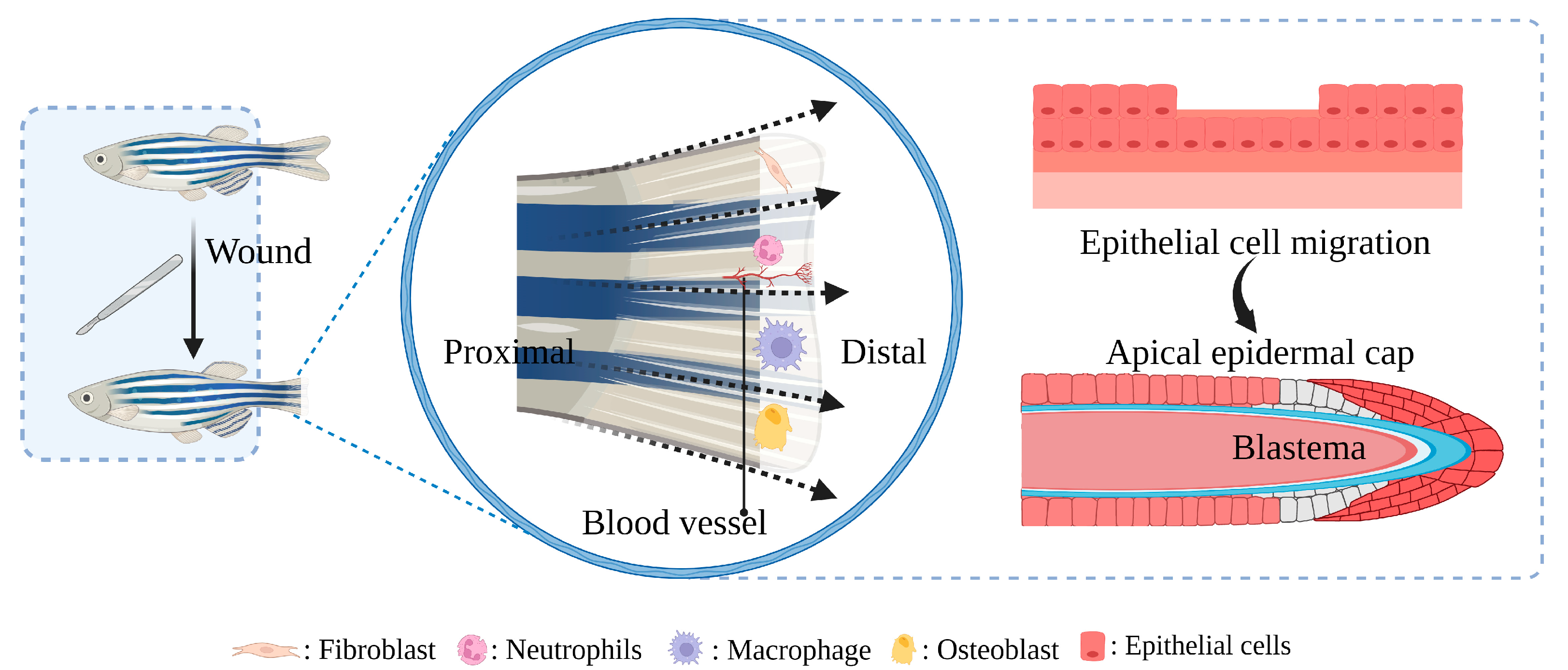
2. Mechanism of Delayed Healing of Diabetic Wounds

2.1. Immune Dysfunction
The inflammatory phase of the wound can last for weeks or even months under the influence of diabetes [54,55][31][32]. The repair process (bacterial defense, cell proliferation, and collagen synthesis) requires energy, and the strictly oxygen-dependent NADPH-linked oxygenase will produce ROS during this period. Many pathways (polyol pathway, hexosamine pathway, etc.) related to oxidative stress of AGEs are maladjusted, resulting in NADPH depletion, decreased glutathione production, activation of protein kinase C and NADPH oxidases, and excessive production of oxygen free radicals and ROS, which further magnify the inflammatory response under high glucose conditions [7,56][7][33]. The functional impairment of neutrophils and macrophages was manifested in the continuous activation of citrulline histone H3, NETosis, and irregular release of NETs in diabetic wounds. Macrophages were affected by increased secretion of prostaglandin E2/D2 and excessive accumulation of AGEs, resulting in weakening of burial and phagocytosis of polymorphonuclear leukocytes such as apoptotic neutrophils, which remained in the M1 phenotype for a long time [17,21,57][20][34][35].
2.2. Microbial Invasion
2.3. Impaired Cell Proliferation and Angiogenesis
In the proliferative stage, the cascade mediated by different matrix metalloproteinases, cytokines, inflammatory cells, keratinocytes, fibroblasts, and endothelial cells is the basis of successful wound healing [62,63,64,65][39][40][41][42]. On the one hand, keratinocyte and fibroblast proliferation and migration provide basic conditions for ECM and re-epithelialization [66,67,68][43][44][45]. On the other hand, a continuous supply of blood provides adequate oxygen and cytokines, which are necessary for tissue regeneration. Wound areas with active proliferating fibroblasts can only be seen when pO2 is above 15 mm Hg [7,23][7][46]. Further, keratinocyte, fibroblast proliferation and migration provide basic conditions for ECM and re-epithelialization of ECM [69][47]. Decreased levels of heat shock proteins (HSPs) (HSP90, HSP70, HSP47, and HSP27) and their downstream molecules TLR4 and p38-mitogen-activated protein kinases affect procollagen synthesis and protein homeostasis [21,70][20][48]. Irregular pro-inflammatory response can activate the irrational upregulation of activating transcription factor-3 and inducible nitric oxide synthase (iNOS), accompanied by the increasing level of free radicals and the upgradulating activities of caspase-3, -8 and -9. High oxidative stress results in the down-regulation of nuclear factor E2-related factor 2 (affecting the expression of MMP-9, transforming growth factor-beta (TGF-β), migration and proliferation-related genes). Impaired cell differentiation and remodeling are associated with abnormal Bcl2, keratin K16, notch junction protein 43, and platelet reactive protein- 1 expression. In addition, the expression of angiopoietin-like 4, one of the stromal cell proteins, is difficult to up-regulate in diabetic wounds as occurs in normal wounds. The activation of the JAK1/STAT3/iNOS signaling pathway is blocked, and the production of NO is inhibited, hinting to angiogenesis and re-epithelialization [21,[20]232.4. Pathological Scar
The remodeling phase requires the synthesis of new collagen and concurrent collagen degradation, a process mediated by MMPs. Type III collagen is gradually replaced by type I collagen with greater tensile strength. The presence of myofibroblasts also causes wound contraction, contributing to potential scar formation [23][46]. However, premature senescence of fibroblasts occurs under high glucose conditions, which is associated with persistent oxidative stress and inflammation [42][18]. At the same time, pathological skin scarring exhibits excessive accumulation of fibroblasts and ECM (mainly type I collagen). The vertical growth of pathological scars generally subsides after a few years; however, the joint site can lead to contracture, severely limiting function, requiring surgical treatment, and seriously affecting appearance and normal life [42][18]. Several lines of evidence suggest that inflammatory bodies in local fibroblasts are involved in skin fibrosis by inducing these normally stationary cells to differentiate into pathogenic myofibroblasts, resulting in high levels of ECM. This inflammasome activation in these non-immune cells triggers skin fibrosis, and subsequent inflammasome activation in immune cells amplifies the local inflammatory response [58][49].3. The Advantages of Zebrafish for Diabetic Wound Healing
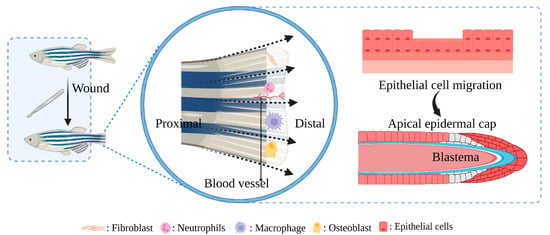
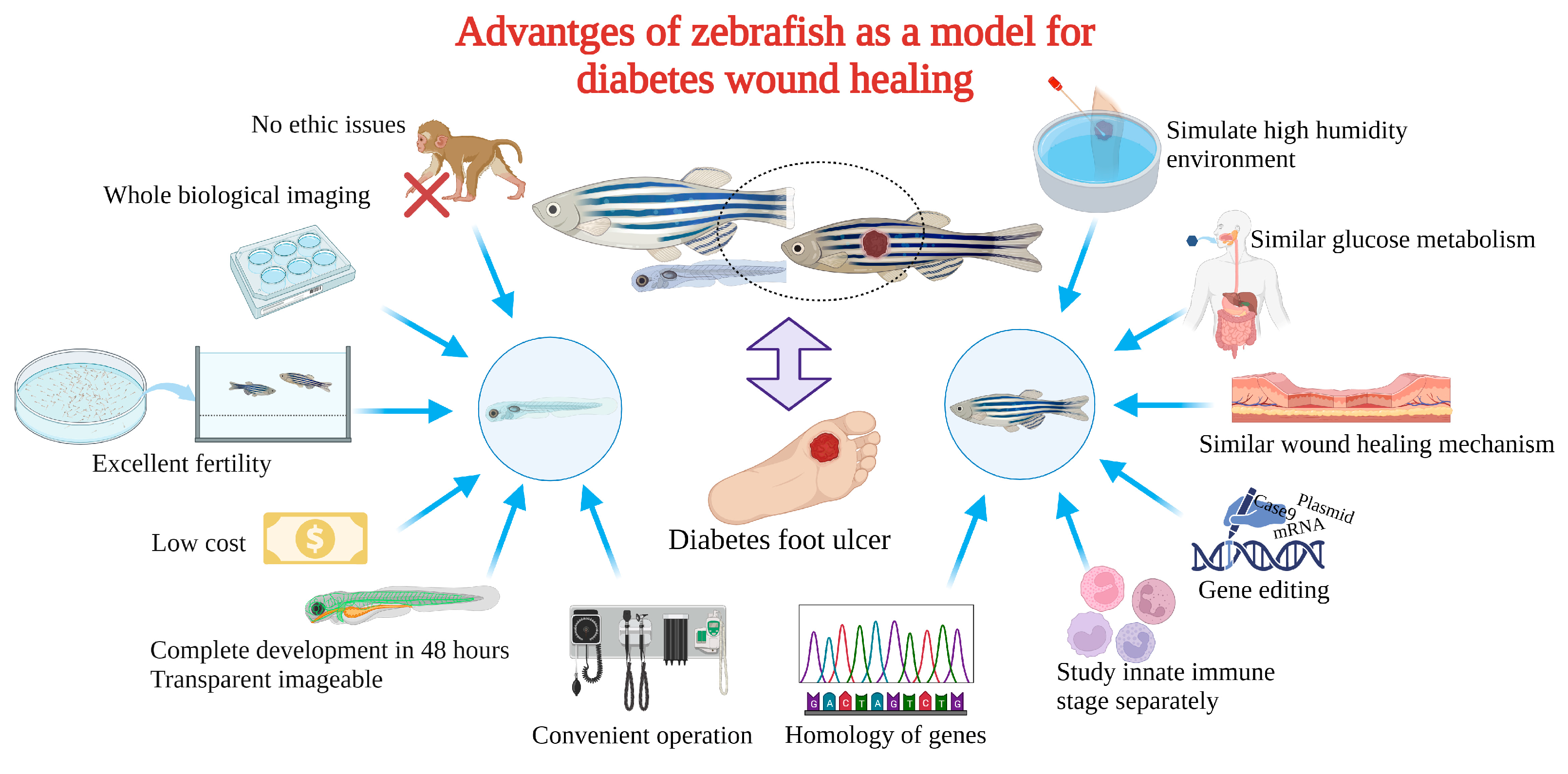
Zebrafish as a model for studying diabetic wound healing has many unique advantages (Figure 3): Zebrafish larvae are only subject to animal experimentation regulations upon reaching 120 h post fertilization and starting to live free, as stipulated by the directive of the European Union on the Protection of Animals used for scientific purposes. As the volume is small, whole-organism imaging can be performed in a multiwell plate; the cost is low; the reproduction ability is strong (about 200 eggs per female per week) [75][50]; the generation time is short, generally 3–4 months. Thus, selection experiments can be conducted; larvae developed rapidly, and fully developed juveniles can be obtained 48 h after hatching. The embryonic and juvenile stages are transparent throughout the body, making real-time imaging of target organs possible, giving ease of embryonic manipulation, and the possibility of screening therapeutic agents at a low cost [76,77,78][51][52][53]. All of the above contribute to their increasing popularity for high-throughput and high-content assays. On the other hand, as a result of the zebrafish genome sequence, 71% of human proteins and 82% of disease-causing human proteins have been found to be orthologous in zebrafish, with significant homology to human proteins. The mechanisms of infection and inflammation caused by innate immune responses can be isolated and studied in the early stages of development without the complications associated with adaptive immune responses. It is suitable for the study of gene expression regulation by gene specific knockout technique or by mRNA or plasmid overexpression of protein. There is good genetic control and in the embryonic and larval stages, the skin of zebrafish is already composed of a superficial peritrind, a middle epidermal double layer, and a basal layer attached to the basal membrane, while its multilayer epidermis is formed during the metamorphosis on the 25th day after fertilization. At the same time, fibroblasts penetrate the dermis, take over the collagen produced by keratinocytes in the basal layer, form locally thickened dermal papillae, and begin to scale. In short, the skin structure of zebrafish is very similar to that of human beings, and the basic principle of the wound healing mechanism is conservative between human and zebrafish: there are discrete stages of healing, allowing specific processes to be studied separately; a remarkable ability to regenerate new fin tissue after amputation is retained, and its caudal fin has a relatively simple but symmetrical structure, including epidermis, blood vessels, nerves, pigment cells, and fibroblasts [39,79,80,81,82,83][54][55][56][57][58][59].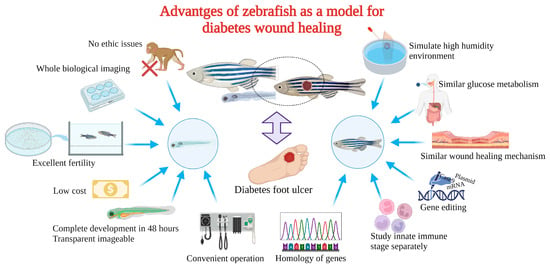
4. Construction and Application of Zebrafish Diabetic Wound Model
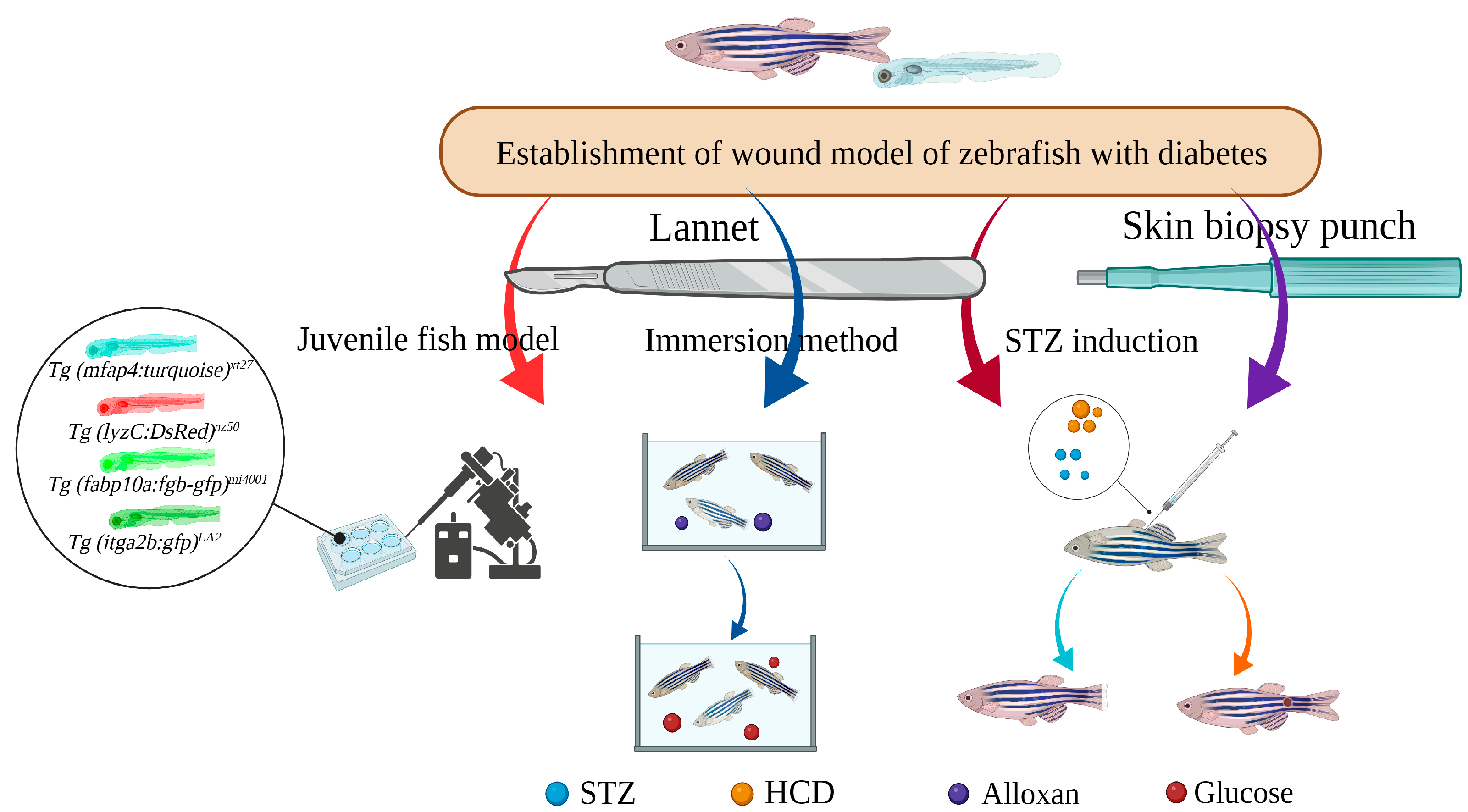
4.1. STZ-Induced Caudal Fin Regeneration Model of Zebrafish with Type 1 Diabetes
4.2. Caudal Fin Model of Type II Diabetes in Adult Zebrafish Induced by Alloxan and Glucose Combined with Aqueous Solution Exposure
4.2. Caudal Fin Model of Type II Diabetes in Adult Zebrafish Induced by Alloxan and Glucose Combined with Aqueous Solution Exposure
Aquaporin (AQP) and GLUT1, both present in the gill and skin epithelium, are thought to be responsible for the production of HG zebrafish following the application of alloxan and glucose in water. Wibowo et al. combined 0.4% Alloxan and glucose (E-Merck) % solution, and placed adult zebrafish aged from 3 to 6 months in 100 mL Alloxan solution for 1 h a day for 5 days, and then transferred 2L 2% glucose solution for 24 h for 6 days, as well as amputating using a lancet (Aesculap Scalpel Handle No. 3 and Aesculap Scalpel Blade No. 10 of B BRAUN) at the first or the second segment below the level of the first ray bifurcation. The expressions of shha, igf2a, bmp2b, and col1a2 were down-regulated in the experimental group, which may be related to the glucose metabolism inducing the generation of superoxide or ROS, the inhibition of GAPDH, the accumulation of methyl glyoxal, and the disruption of hypoxia inducible factor-1α transcription factor stability, leading to the transcriptional inhibition of some of the above target genes. The percentage of caudal fin regeneration and the expression of shha, igf2a, bmp2b, and col1a2 were increased after treatment with 15 ppm propolis ethanol extract [98][64].4.3. Caudal Fin Regeneration Model of Zebrafish Juvenile Type II Diabetes Induced by Single Immersion or Injection of Glucose
4.4. Skin Wound Model of Adult Zebrafish Type I Diabetes Induced by STZ Injection
The full-layer wound healing mechanism of zebrafish is very similar to that of humans. On the one hand, in the embryonic and larval stages, the skin of zebrafish is already composed of the outer layer of peritrind, the middle layer of epidermis, and the basal layer attached to the basal membrane. In the process of metamorphosis on the 25th day after fertilization, multiple layers of epidermis are formed. At the same time, fibroblasts penetrate the dermis and take over collagen produced by basal keratinocytes to form locally thickened dermal papilla and scale, which is very similar to human skin structure. On the other hand, skin healing of zebrafish involves activation of signal transduction pathways downstream of hydrogen peroxide, including epidermal growth factor EGF, forkhead box-1, and IkappaB kinase-alpha. EGF regulates TGF-β through ERK1/2 and EGFR signal transduction. Damaged cells bind to EGFR to trigger cascades (such as TGF-β/integrin and ROCK/JNK pathways) to induce DNA synthesis and cell proliferation at the wound site.5. Conclusions
Delayed wound healing induced by diabetes results in a heavy burden on patients physically, mentally, and economically. To discover specific drugs with excellent efficacy, it is essential to investigate the specific mechanism behind the delayed feature utilizing appropriate preclinical research models. Zebrafish are characterized by high throughput, small ethical disputes, larval body transparency, quick development, fast and efficient establishment of a model of diabetes, convenient caudal fin regeneration of wound repair evaluation, and are the height of human gene homology, etc. In addition, quantitative gavage and feeding shrimp with high glucose can solve restrictions such as water-soluble exposure and potential damage by injection. Zebrafish will become an ideal model for diabetic wound model organisms in the future, bringing a new dawn for the screening of rapid healing treatment for diabetic wounds and the exploration of the underlying mechanism.References
- Chen, W.; Mao, M.; Fang, J.; Xie, Y.; Rui, Y. Fracture Risk Assessment in Diabetes Mellitus. Front. Endocrinol. 2022, 13, 961761.
- Tsunoda, T.; Samadi, A.; Burade, S.; Mahmud, T. Complete Biosynthetic Pathway to the Antidiabetic Drug Acarbose. Nat. Commun. 2022, 13, 3455.
- Giugliano, D.; Longo, M.; Caruso, P.; Di Fraia, R.; Scappaticcio, L.; Gicchino, M.; Petrizzo, M.; Bellastella, G.; Maiorino, M.I.; Esposito, K. Feasibility of Simplification From a Basal-Bolus Insulin Regimen to a Fixed-Ratio Formulation of Basal Insulin Plus a GLP-1RA or to Basal Insulin Plus an SGLT2 Inhibitor: BEYOND, a Randomized, Pragmatic Trial. Diabetes Care 2021, 44, 1353–1360.
- Li, S.; Mohamedi, A.H.; Senkowsky, J.; Nair, A.; Tang, L. Imaging in Chronic Wound Diagnostics. Adv. Wound Care New Rochelle 2020, 9, 245–263.
- Velnar, T.; Bailey, T.; Smrkolj, V. The Wound Healing Process: An Overview of the Cellular and Molecular Mechanisms. J. Int. Med. Res. 2009, 37, 1528–1542.
- Wilkinson, H.N.; Hardman, M.J. Wound Healing: Cellular Mechanisms and Pathological Outcomes. Open Biol. 2020, 10, 200223.
- Schreml, S.; Szeimies, R.M.; Prantl, L.; Karrer, S.; Landthaler, M.; Babilas, P. Oxygen in Acute and Chronic Wound Healing: Oxygen in Wound Healing. Br. J. Dermatol. 2010, 163, 257–268.
- Holl, J.; Kowalewski, C.; Zimek, Z.; Fiedor, P.; Kaminski, A.; Oldak, T.; Moniuszko, M.; Eljaszewicz, A. Chronic Diabetic Wounds and Their Treatment with Skin Substitutes. Cells 2021, 10, 655.
- Zheng, Z.; Wan, Y.; Liu, Y.; Yang, Y.; Tang, J.; Huang, W.; Cheng, B. Sympathetic Denervation Accelerates Wound Contraction but Inhibits Reepithelialization and Pericyte Proliferation in Diabetic Mice. J. Diabetes Res. 2017, 2017, 7614685.
- Randi, A.M.; Laffan, M.A. Von Willebrand Factor and Angiogenesis: Basic and Applied Issues. J. Thromb. Haemost. 2017, 15, 13–20.
- Zindle, J.K.; Wolinsky, E.; Bogie, K.M. A Review of Animal Models from 2015 to 2020 for Preclinical Chronic Wounds Relevant to Human Health. J. Tissue Viability 2021, 30, 291–300.
- Chen, C.; Gu, Y.; Philippe, J.; Zhang, P.; Bachman, H.; Zhang, J.; Mai, J.; Rufo, J.; Rawls, J.F.; Davis, E.E.; et al. Acoustofluidic Rotational Tweezing Enables High-Speed Contactless Morphological Phenotyping of Zebrafish Larvae. Nat. Commun. 2021, 12, 1118.
- Leung, T.; Chen, H.; Stauffer, A.M.; Giger, K.E.; Sinha, S.; Horstick, E.J.; Humbert, J.E.; Hansen, C.A.; Robishaw, J.D. Zebrafish G Protein Gamma2 Is Required for VEGF Signaling during Angiogenesis. Blood 2006, 108, 160–166.
- Li, Y.-W.; Chiang, K.-Y.; Li, Y.-H.; Wu, S.-Y.; Liu, W.; Lin, C.-R.; Wu, J.-L. MiR-145 Mediates Zebrafish Hepatic Outgrowth through Progranulin A Signaling. PLoS ONE 2017, 12, e0177887.
- Zhao, S.; Xia, J.; Wu, X.; Zhang, L.; Wang, P.; Wang, H.; Li, H.; Wang, X.; Chen, Y.; Agnetti, J.; et al. Deficiency in Class III PI3-Kinase Confers Postnatal Lethality with IBD-like Features in Zebrafish. Nat. Commun. 2018, 9, 2639.
- Wilkinson, R.N.; Koudijs, M.J.; Patient, R.K.; Ingham, P.W.; Schulte-Merker, S.; van Eeden, F.J.M. Hedgehog Signalling via a Calcitonin Receptor-like Receptor Can Induce Arterial Differentiation Independently of VEGF Signalling in Zebrafish. Blood 2012, 120, 477–488.
- Berger, J.; Li, M.; Berger, S.; Meilak, M.; Rientjes, J.; Currie, P.D. Effect of Ataluren on Dystrophin Mutations. J. Cell Mol. Med. 2020, 24, 6680–6689.
- Eming, S.A.; Martin, P.; Tomic-Canic, M. Wound Repair and Regeneration: Mechanisms, Signaling, and Translation. Sci. Transl. Med. 2014, 6, 265sr6.
- Wang, Y.; Chen, L.; Ren, D.-Y.; Feng, Z.-X.; Zhang, L.-Y.; Zhong, Y.-F.; Jin, M.-Y.; Xu, F.-W.; Feng, C.-Y.; Du, Y.-Z.; et al. Mussel-Inspired Collagen-Hyaluronic Acid Composite Scaffold with Excellent Antioxidant Properties and Sustained Release of a Growth Factor for Enhancing Diabetic Wound Healing. Mater. Today Bio 2022, 15, 100320.
- Patel, S.; Srivastava, S.; Singh, M.R.; Singh, D. Mechanistic Insight into Diabetic Wounds: Pathogenesis, Molecular Targets and Treatment Strategies to Pace Wound Healing. Biomed. Pharmacother. 2019, 112, 108615.
- Wang, D.; Chen, H.; Lei, L.; Chen, J.; Gao, J.; Liu, J.; Li, Q.; Xie, Y.; Hu, Y.; Ni, Y. Biofabricated Macrophage and Fibroblast Membranes Synergistically Promote Skin Wound Healing. Bioeng. Transl. Med. 2022, 7, e10344.
- Ma, J.; Yong, L.; Lei, P.; Li, H.; Fang, Y.; Wang, L.; Chen, H.; Zhou, Q.; Wu, W.; Jin, L.; et al. Advances in MicroRNA from Adipose-Derived Mesenchymal Stem Cell-Derived Exosome: Focusing on Wound Healing. J. Mater. Chem. B 2022, 10, 9565–9577.
- Liu, J.; Carnero-Montoro, E.; van Dongen, J.; Lent, S.; Nedeljkovic, I.; Ligthart, S.; Tsai, P.-C.; Martin, T.C.; Mandaviya, P.R.; Jansen, R.; et al. An Integrative Cross-Omics Analysis of DNA Methylation Sites of Glucose and Insulin Homeostasis. Nat. Commun. 2019, 10, 2581.
- Beserra, F.P.; Vieira, A.J.; Gushiken, L.F.S.; de Souza, E.O.; Hussni, M.F.; Hussni, C.A.; Nóbrega, R.H.; Martinez, E.R.M.; Jackson, C.J.; de Azevedo Maia, G.L.; et al. Lupeol, a Dietary Triterpene, Enhances Wound Healing in Streptozotocin-Induced Hyperglycemic Rats with Modulatory Effects on Inflammation, Oxidative Stress, and Angiogenesis. Oxid. Med. Cell Longev. 2019, 2019, 3182627.
- Feng, W.; Zhang, C.; Yu, T.; Zhu, D. Quantitative Evaluation of Skin Disorders in Type 1 Diabetic Mice by in vivo Optical Imaging. Biomed. Opt. Express 2019, 10, 2996–3008.
- Makvandi, P.; Caccavale, C.; Della Sala, F.; Zeppetelli, S.; Veneziano, R.; Borzacchiello, A. Natural Formulations Provide Antioxidant Complement to Hyaluronic Acid-Based Topical Applications Used in Wound Healing. Polymers 2020, 12, 1847.
- Brazel, C.B.; Simon, J.C.; Tuckermann, J.P.; Saalbach, A. Inhibition of 11β-HSD1 Expression by Insulin in Skin: Impact for Diabetic Wound Healing. J. Clin. Med. 2020, 9, 3878.
- Rozman, N.A.S.; Tong, W.Y.; Leong, C.R.; Anuar, M.R.; Karim, S.; Ong, S.K.; Yusof, F.A.M.; Tan, W.-N.; Sulaiman, B.; Ooi, M.L.; et al. Homalomena Pineodora Essential Oil Nanoparticle Inhibits Diabetic Wound Pathogens. Sci. Rep. 2020, 10, 3307.
- Augustine, R.; Zahid, A.A.; Hasan, A.; Wang, M.; Webster, T.J. CTGF Loaded Electrospun Dual Porous Core-Shell Membrane for Diabetic Wound Healing. Int. J. Nanomed. 2019, 14, 8573–8588.
- Ma, J.; Lei, P.; Chen, H.; Wang, L.; Fang, Y.; Yan, X.; Yang, Q.; Peng, B.; Jin, L.; Sun, D. Advances in LncRNAs from Stem Cell-Derived Exosome for the Treatment of Cardiovascular Diseases. Front. Pharmacol. 2022, 13, 986683.
- Jiang, T.; Liu, S.; Wu, Z.; Li, Q.; Ren, S.; Chen, J.; Xu, X.; Wang, C.; Lu, C.; Yang, X.; et al. Smart Hydrogel Promotes Diabetic Wound Healing by Optimizing Cellular Functions and Relieving Oxidative Stress. Mater. Today Bio 2022, 16, 100365.
- Tilves, C.M.; Zmuda, J.M.; Kuipers, A.L.; Nestlerode, C.S.; Evans, R.W.; Bunker, C.H.; Patrick, A.L.; Miljkovic, I. Association of Lipopolysaccharide-Binding Protein with Aging-Related Adiposity Change and Prediabetes Among African Ancestry Men. Diabetes Care 2016, 39, 385.
- Ji, H.; Peng, R.; Jin, L.; Ma, J.; Yang, Q.; Sun, D.; Wu, W. Recent Advances in ROS-Sensitive Nano-Formulations for Atherosclerosis Applications. Pharmaceutics 2021, 13, 1452.
- Eichelberger, K.R.; Goldman, W.E. Manipulating Neutrophil Degranulation as a Bacterial Virulence Strategy. PLoS Pathog. 2020, 16, e1009054.
- Zhang, J.; Dai, Y.; Wei, C.; Zhao, X.; Zhou, Q.; Xie, L. DNase I Improves Corneal Epithelial and Nerve Regeneration in Diabetic Mice. J. Cell Mol. Med. 2020, 24, 4547–4556.
- Moura, J.; Rodrigues, J.; Gonçalves, M.; Amaral, C.; Lima, M.; Carvalho, E. Impaired T-Cell Differentiation in Diabetic Foot Ulceration. Cell Mol. Immunol. 2017, 14, 758–769.
- Daeschlein, G. Antimicrobial and Antiseptic Strategies in Wound Management. Int. Wound J. 2013, 10 (Suppl. 1), 9–14.
- Tang, N.; Zheng, Y.; Jiang, X.; Zhou, C.; Jin, H.; Jin, K.; Wu, W.; Haick, H. Wearable Sensors and Systems for Wound Healing-Related PH and Temperature Detection. Micromachines 2021, 12, 430.
- Pang, J.; Urao, N.; Koh, T.J. Proliferation of Ly6C+ Monocytes/Macrophages Contributes to Their Accumulation in Mouse Skin Wounds. J. Leukoc. Biol. 2020, 107, 551–560.
- Icli, B.; Nabzdyk, C.S.; Lujan-Hernandez, J.; Cahill, M.; Auster, M.E.; Wara, A.; Sun, X.; Ozdemir, D.; Giatsidis, G.; Orgill, D.P.; et al. Regulation of Impaired Angiogenesis in Diabetic Dermal Wound Healing by MicroRNA-26a. J. Mol. Cell Cardiol. 2016, 91, 151–159.
- Chen, C.-Y.; Yin, H.; Chen, X.; Chen, T.-H.; Liu, H.-M.; Rao, S.-S.; Tan, Y.-J.; Qian, Y.-X.; Liu, Y.-W.; Hu, X.-K.; et al. Ångstrom-Scale Silver Particle–Embedded Carbomer Gel Promotes Wound Healing by Inhibiting Bacterial Colonization and Inflammation. Sci. Adv. 2020, 6, eaba0942.
- Chen, C.-Y.; Rao, S.-S.; Ren, L.; Hu, X.-K.; Tan, Y.-J.; Hu, Y.; Luo, J.; Liu, Y.-W.; Yin, H.; Huang, J.; et al. Exosomal DMBT1 from Human Urine-Derived Stem Cells Facilitates Diabetic Wound Repair by Promoting Angiogenesis. Theranostics 2018, 8, 1607–1623.
- Seo, G.Y.; Ho, M.T.; Bui, N.T.; Kim, Y.M.; Koh, D.; Lim, Y.; Hyun, C.; Cho, M. Novel Naphthochalcone Derivative Accelerate Dermal Wound Healing through Induction of Epithelial-Mesenchymal Transition of Keratinocyte. J. Biomed. Sci. 2015, 22, 47.
- Zhang, Z.; Zi, Z.; Lee, E.E.; Zhao, J.; Contreras, D.C.; South, A.P.; Abel, E.D.; Chong, B.F.; Vandergriff, T.; Hosler, G.A.; et al. Differential Glucose Requirement in Skin Homeostasis and Injury Identifies a Therapeutic Target for Psoriasis. Nat. Med. 2018, 24, 617–627.
- Pedowitz, N.J.; Batt, A.R.; Darabedian, N.; Pratt, M.R. MYPT1 O-GlcNAc Modification Regulates Sphingosine-1-Phosphate Mediated Contraction. Nat. Chem. Biol. 2021, 17, 169–177.
- Hu, S.C.-S.; Lan, C.-C.E. High-Glucose Environment Disturbs the Physiologic Functions of Keratinocytes: Focusing on Diabetic Wound Healing. J. Dermatol. Sci. 2016, 84, 121–127.
- Nishikori, Y.; Shiota, N.; Okunishi, H. The Role of Mast Cells in Cutaneous Wound Healing in Streptozotocin-Induced Diabetic Mice. Arch. Dermatol. Res. 2014, 306, 823–835.
- Park, K.H.; Han, S.H.; Hong, J.P.; Han, S.-K.; Lee, D.-H.; Kim, B.S.; Ahn, J.H.; Lee, J.W. Topical Epidermal Growth Factor Spray for the Treatment of Chronic Diabetic Foot Ulcers: A Phase III Multicenter, Double-Blind, Randomized, Placebo-Controlled Trial. Diabetes Res. Clin. Pract. 2018, 142, 335–344.
- Huang, C.; Ogawa, R. Role of Inflammasomes in Keloids and Hypertrophic Scars—Lessons Learned from Chronic Diabetic Wounds and Skin Fibrosis. Int. J. Mol. Sci. 2022, 23, 6820.
- Maierdiyali, A.; Wang, L.; Luo, Y.; Li, Z. Effect of Tank Size on Zebrafish Behavior and Physiology. Animals 2020, 10, 2353.
- Andersson, O.; Adams, B.A.; Yoo, D.; Ellis, G.C.; Gut, P.; Anderson, R.M.; German, M.S.; Stainier, D.Y.R. Adenosine Signaling Promotes Regeneration of Pancreatic β-Cells in vivo. Cell Metab. 2012, 15, 885–894.
- Moustaqil, M.; Fontaine, F.; Overman, J.; McCann, A.; Bailey, T.L.; Soto, P.R.; Bhumkar, A.; Giles, N.; Hunter, D.J.B.; Gambin, Y.; et al. Homodimerization Regulates an Endothelial Specific Signature of the SOX18 Transcription Factor. Nucleic Acids Res. 2018, 46, 11381–11395.
- Soman, S.; Keatinge, M.; Moein, M.; Da Costa, M.; Mortiboys, H.; Skupin, A.; Sugunan, S.; Bazala, M.; Kuznicki, J.; Bandmann, O. Inhibition of the Mitochondrial Calcium Uniporter Rescues Dopaminergic Neurons in pink1−/− Zebrafish. Eur. J. Neurosci. 2017, 45, 528–535.
- Richardson, R.; Slanchev, K.; Kraus, C.; Knyphausen, P.; Eming, S.; Hammerschmidt, M. Adult Zebrafish as a Model System for Cutaneous Wound-Healing Research. J. Investig. Dermatol. 2013, 133, 1655–1665.
- LeBert, D.C.; Huttenlocher, A. Inflammation and Wound Repair. Semin. Immunol. 2014, 26, 315–320.
- Jörgens, K.; Hillebrands, J.-L.; Hammes, H.-P.; Kroll, J. Zebrafish: A Model for Understanding Diabetic Complications. Exp. Clin. Endocrinol. Diabetes 2012, 120, 186–187.
- Martínez-Navarro, F.J.; Martínez-Morcillo, F.J.; de Oliveira, S.; Candel, S.; Cabas, I.; García-Ayala, A.; Martínez-Menchón, T.; Corbalán-Vélez, R.; Mesa-del-Castillo, P.; Cayuela, M.L.; et al. Hydrogen Peroxide in Neutrophil Inflammation: Lesson from the Zebrafish. Dev. Comp. Immunol. 2020, 105, 103583.
- Yamamoto, D.; Sato, D.; Nakayama, H.; Nakagawa, Y.; Shimada, Y. ZF-Mapper: Simple and Complete Freeware for Fluorescence Quantification in Zebrafish Images. Zebrafish 2019, 16, 233–239.
- Naomi, R.; Bahari, H.; Yazid, M.D.; Embong, H.; Othman, F. Zebrafish as a Model System to Study the Mechanism of Cutaneous Wound Healing and Drug Discovery: Advantages and Challenges. Pharmaceuticals 2021, 14, 1058.
- Cho, K.-H.; Kim, J.-H.; Nam, H.-S.; Kang, D.-J. Efficacy Comparison Study of Human Epidermal Growth Factor (EGF) between Heberprot-P® and Easyef® in Adult Zebrafish and Embryo under Presence or Absence Combination of Diabetic Condition and Hyperlipidemia to Mimic Elderly Patients. Geriatrics 2022, 7, 45.
- Yoon, J.-H.; Cho, K.-H. A Point Mutant of Apolipoprotein A-I (V156K) Showed Enhancement of Cellular Insulin Secretion and Potent Activity of Facultative Regeneration in Zebrafish. Rejuvenation Res. 2012, 15, 313–321.
- Intine, R.V.; Olsen, A.S.; Sarras, M.P. A Zebrafish Model of Diabetes Mellitus and Metabolic Memory. J. Vis. Exp. 2013, 72, e50232.
- Sarras, M.P.; Leontovich, A.A.; Olsen, A.S.; Intine, R.V. Impaired Tissue Regeneration Corresponds with Altered Expression of Developmental Genes That Persists in the Metabolic Memory State of Diabetic Zebrafish. Wound Repair Regen 2013, 21, 320–328.
- Wibowo, I.; Utami, N.; Anggraeni, T.; Barlian, A.; Putra, R.E.; Indriani, A.D.; Masadah, R.; Ekawardhani, S. Propolis Can Improve Caudal Fin Regeneration in Zebrafish (Danio rerio) Induced by The Combined Administration of Alloxan and Glucose. Zebrafish 2021, 18, 274–281.
- Morris, S.; Cholan, P.M.; Britton, W.J.; Oehlers, S.H. Glucose Inhibits Haemostasis and Accelerates Diet-Induced Hyperlipidaemia in Zebrafish Larvae. Sci. Rep. 2021, 11, 19049.