2. Epigenetic Regulation of α-Synuclein
DNAm, which usually includes the addition of a methyl group to one of the carbons of the pyrimidine ring of cytosine in the CpG dinucleotide, has been broadly characterized in human individuals. In addition to its plasticity towards genes and the environment, DNAm also has a plausible application as a biomarker because of the similarity of DNAm patterns between the blood and brain
[8].
α-Synuclein expression is regulated by multiple mechanisms, one of which is the methylation of intron 1 region [SNCA(i1)] in α-synuclein gene located upstream of the initiation codon ATG, between exon 1 and exon 2. This region contains CpG islands which serve as binding sites for several transcription factors. Binding of transcription factors to these binding sites regulates α-synuclein expression
[2][9][2,9]. Hypomethylation in this region increases α-synuclein expression which may cause its accumulation and ultimately lead to pathology
[10].
Daniele et al. (2018)
[11] demonstrated that the SNCA(i1) methylation level was augmented with the age of an individual, reducing α-synuclein concentration in the blood. Physical activity possessed a similar effect, decreasing both the oligomeric and total α-synuclein levels. The authors concluded that DNAm of SNCA(i1) was associated with age, and total α-synuclein concentrations in erythrocytes were decreased with age. DNA methyltransferase (DNMT) content in this
res
earchtudy was in good correlation with oligomeric and total α-synuclein, while total α-synuclein concentration was inversely related to the methylation status of SNCA(i1).
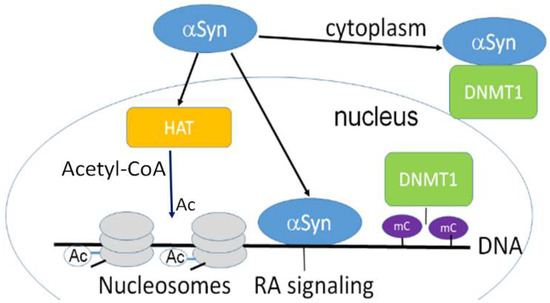
α-Synuclein can retain DNMT1 in the cytoplasmic fraction of neurons (
Figure 1) affecting DNAm. As a result, reduced methylation was detected in SNCA(i1)
[12].
Figure 1. α-Synuclein is confined both in the nucleus and cytoplasm and is involved in epigenetic regulation. Nuclear α-synuclein binding to DNA regulates specific gene transcription, reducing histone acetyltransferase p300 (HAT) activity. α-Synuclein is able to sequester DNMT1 from the nucleus to the cytoplasm. HAT acetylates conserved lysine residues on histones. This reaction includes the transfer of an acetyl group (Ac) from acetyl-CoA to form ε-N-acetyl-lysine. Histone acetylation switches genes on and off, acting as a regulator of gene expression. DNA is wrapped around histones forming nucleosomes (gray disks). Nuclear α -synuclein also participates in the retinoic acid (RA) regulation of the signaling pathway. mC-CpG methylation.
Several factors, including variations in the α-synuclein gene, physical activity of an individual, exposure to pesticides, etc. may impact the epigenome, particularly at the level of CpG methylation
[5][11][12][5,11,12]. These factors are key contributors to the pathology of synucleinopathies. The results of many recent studies demonstrate the importance of the interaction between genetic and environmental contributions to the disease risk, presumably acting across multiple omics layers
[5][13][5,13].
The methylation level of SNCA(i1) is different in Parkinson’s disease patients and control individuals in blood, leukocytes, and mononuclear cells. The results of recent investigations demonstrate that the level of methylation in patients is considerately lower than in a control group. A similar conclusion was made when brain tissue methylation was analyzed
[14][15][16][14,15,16]. Thus, complex interactions between genotype, environment, lifestyle, and developmental stage affect and regulate epigenetic modifications
[17][18][17,18].
3. α-Synuclein as a Genetic Modulator
3.1. Nuclear Localization of α-Synuclein and DNA Binding
After finding α-synuclein in the nucleus and presynaptic nerve terminals
[19], the association of neuritic and nuclear α-synuclein, and the functional activity and interacting partners of α-synuclein in subcellular compartments, have been the focus of attention of many investigations. However, the biology of nuclear α-synuclein is still not completely apprehended.
The interactions of both wild-type and mutant forms of α-synuclein with dsDNA were described for the first time by Cherny et al. (2004)
[20], and later confirmed by several studies
[21][22][23][21,22,23]. α-Synuclein-DNA binding alters the properties of both interacting molecules. α-Synuclein–DNA interaction changes DNA bending properties and increases aggregation of the protein, with a concomitant fibril formation. Although, at present, a clear answer to the meaning of this interaction is under discussion, an interesting hypothesis is that α-synuclein binds to histone-free segments of DNA which are transcriptionally active, and that this interaction causes a transcription activity alteration
[20]. Binding of α-synuclein to specific nucleotide sequences in DNA has been later repeatedly confirmed
[23][24][25][26][23,24,25,26].
α-Synuclein binding to DNA modulates several DNA- and RNA-dependent activities, including transcription, rRNA and mRNA metabolism
[25][27][28][25,27,28], and DNA double-strand break repair
[24]. Moreover, α-synuclein induces DNA stretches and increases its stiffness
[29]. However, probably the most exciting and stimulating were the studies investigating the involvement of α-synuclein in epigenetic mechanisms, for example its role in the regulation of histone modification
[30].
The biochemical functions of nuclear α-synuclein demonstrate that it binds both to DNA and histones. This interaction is physiologically important because it controls DNA repair and transcriptional regulation
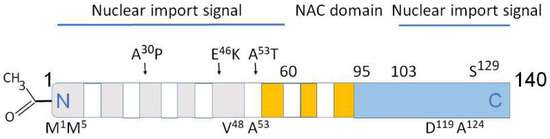
[24][31][24,31]. Interestingly, during neuronal differentiation, distinct α-synuclein species translocate from the nucleus to neuronal processes. α-Synuclein nuclear–somatic–neuritic shuttling is a dynamic process during neuronal differentiation. Nuclear α-synuclein induces noticeable transcriptional deregulation, for example, the downregulation of key cell cycle-related genes. This effect on transcription is related to the reduced α-synuclein binding to DNA. Increased nuclear presence of high-molecular weight α-synuclein regulates specific gene expression. The effect of nuclear α-synuclein on gene expression and cytotoxicity is modulated by phosphorylation on serine 129 (Ser
129,
Figure 2)
[31]. Thus, the activity of nuclear α-synuclein is modulated by the level of its aggregation and post-translational modifications.
Figure 2. Structural domains in α-synuclein involved in aggregation, metal binding and nuclear transport. Above the rectangle: A30P, E46K, A53T—some of mutations causing PD. Below the rectangle: M1, M5 V48, A53—high affinity Cu binding sites. The predominantly disordered C-terminal domain modulates interactions of pro-oxidant metals associated with PD pathology. D119, A124—divalent metals Fe, Cu, Co, and Mn binding sites. Amino acids 1–60 and 103–140 are important for nuclear transport (nuclear import signals). The core sections of the seven amino-terminal repeats (KTKEGV motive) are shown as white bars. These motives are located between amino acids 7–87. Five positively charged regions are light gray, three negatively charged hydrophobic regions are orange, C-terminal region is light blue. Non-amyloid-β component (NAC domain) constituting amino acids 61–95 is critical for aggregation. N-terminal acetylation (left) slows down α-synuclein aggregation and changes the morphology of the aggregates.
Pinho and co-authors
[31] investigated the activity of aggregated and phosphorylated α-synuclein in the nucleus, comparing the results of studies in postmortem brain samples and in different cell models. The authors found that phosphorylation of α-synuclein modulated both its nuclear localization and its role as a regulator of transcription. The results of the epitope dot blot analysis suggest that different conformational states of nuclear and cytosolic α-synuclein with exposed different epitopes determine its interaction with other compounds in the cytoplasm and nucleus
[31].
α-Synuclein also interacts with a protein BAP1 (BRCA1 associated protein 1) possessing ubiquitin C-terminal hydrolase activity (UCH)
[32][38]. BAP1, possessing deubiquitinase activity (DUBs), is required for the reverse reaction of ubiquitination, acting as a major regulator of ubiquitin signaling processes. BAP1 deubiquitinating enzymatic activity is an important modulator of gene expression. The interaction of α-synuclein with BAP1 and other DUBs is a prognostic indicator of unfavorable outcome in several types of cancer, and may also affect neurodegenerative processes
[32][38]. Further studies are necessary to better understand the regulatory role of α-synuclein-BAP1 interaction and its effect on specific gene regulation.
3.2. Nuclear α-Synuclein Regulates Histone Modifications
α-Synuclein interactions with nuclear components are not restricted to DNA. α-Synuclein binds to histones, and its fibrillation is accelerated by histone H1 and other core histones. Co-localization of α-synuclein with histone H1 and H3 and formation of a complex in the nucleus is proven by immunohistochemistry data using neuronal nuclei marker NeuN
[33][39]. The condensation of histones is basically responsible for the organization of euchromatin and heterochromatin, the first being active and the latter transcriptionally repressed. The activity of proteins performing transcription is regulated by various biochemical modifications, i.e., methylation, phosphorylation, and acetylation, on DNA itself or on histones. Post-translational modifications of histone tails may change the protein surface charge, altering its affinity for DNA and for other histones. This, in turn, allows or restricts the entrance of transcription factors and other transcription-associated proteins. Epigenetic “writers”—the proteins responsible for writing these marks—are able to add modifications to histones or DNA. This group includes DNMTs, HAT, and histone methyltransferases (HMTs). These proteins function on the chromatin and make dynamic modifications in response to the environmental factors. Corresponding “erasers”, e.g., histone deacetylases (HDACs), remove alterations to histones or DNA that were introduced by epigenetic writers.
α-Synuclein may interact with epigenetic writers. For example, expressing α-synuclein raises histone-H3 lysine-9 (H3K9) methylation
[34][40]. Overexpressed α-synuclein can retain DNMT1 in the neuronal cytoplasm, changing DNAm (
Figure 1). Moreover, inducible α-synuclein expression increases the level of mono- and dimethylation. These changes may cause the release of synaptic vesicles, contributing to the synaptic dysfunction that occurs in PD
[34][40].
Another important mechanism of α-synuclein’s regulatory effect on transcription is via its interaction with epigenetic erasers. Extensive evidence has demonstrated that α-synuclein restricts HDACs in the cytoplasm, constraining its normal function. Experiments with mutant forms of α-synuclein, e.g., pA
30P and pA
53T, showed that it bound to histones, which in turn decreased histone acetylation (
Figure 1). Thus, HDACs inhibition protects against α-synuclein’s toxic effect, as found in several model systems such as SH-SY5Y cells and transgenic flies
[30].
α-Synuclein interacts with chief epigenetic eraser HDAC4, abundant in neurons. It forms a part of the HDACs class IIa that can shuttle between the nucleus and cytoplasm
[35][41]. Exposition to MPTP (1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine) causes accumulation of HDAC4 in the nucleus. Moreover, nuclear HDAC4 mediates the cell death in A
53T cells by inhibiting CREB (cAMP response element binding protein) and MEF2A (myocyte enhancer factor 2A)
[36][42].
Finally, it has been reported on H4 cells that the pS
129-α-synuclein in the nucleus downregulates important cell-cycle genes, e.g., G2/mitotic-specific cyclin-B1 (CCNB1)—a protein involved in mitosis and a transcription factor modulating the cell cycle (E2F8)
[31].
Furthermore, Schaser et al.
[24] described a new α-synuclein role in the nucleus. A team of researchers demonstrated that α-synuclein phosphorylated on serine-129 (pS
129) rapidly translocated to laser-induced DNA damage sites in the nucleus of in vivo mouse brain cells, as well as in a mouse primary cortical neuron system. This α-synuclein recruitment had a conceivable role in the DNA break repair
[24]. This pathological form is present in the nucleus of neurons in brain areas of mice with impaired cognitive behavioral phenotypes. α-Synuclein’s role in the interaction and modification of histones is essential because histones are key regulators of transcription, and α-synuclein binding alters histones regulatory functions.