Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Isabella Reccia and Version 3 by Conner Chen.
Tumour heterogeneity is a common phenomenon in gastroenteropancreatic (GEP)-neoplasms (NENs)-NENs and has a negative impact on treatment success and prognosis as it produces cell clones that do not express treatment targets (i.e., SSTR, mammalian target of rapamycin–mTOR- signalling pathway, Ki-67).
- neuroendocrine tumour
- neuroendocrine neoplasms
- gastroenteropancreatic
1. Introduction
Tumour heterogeneity refers to spatial and temporal variations that may occur within the tumour environment (intra-tumour) or within individual tumour foci, and also between tumour sites (inter-tumour) [1]. Such heterogeneity may encompass genetic and epigenetic variations, or differences in the tumour microenvironment [2][3][4][5][6][2,3,4,5,6]. Tumour heterogeneity can also evolve over time due to selective pressures, such as those imposed by treatment, leading to selection and clonal expansion of subpopulations [1][7][8][9][1,7,8,9]. Tumour heterogeneity is common in human tumours and its occurrence is essential to understand and predict tumour progression and response to specific treatment [10]. Higher intra-tumour and/or inter-tumour heterogeneity can be associated with negative outcomes [7].
Neuroendocrine tumours (NETs), better defined as neoplasms (NENs), are a heterogeneous group of neoplasms that range from well-differentiated tumours to more aggressive carcinomas (Table 1) [11].
Table 1.
Classification for gastroenteropancreatic neuroendocrine neoplasms (GEP-NENs).
| Terminology | Differentiation | Grade | Mitotic Rate (Mitoses/2 mm2) * | Ki-67 Index % ** |
|---|---|---|---|---|
| NET, G1 | Well differentiated | Low | <2 | <3% |
| NET, G2 | Intermediate | 2–20 | 3–20% | |
| NET, G3 | High | >20 | >20% | |
| NEC, small-cell type (SCNEC) | Poorly differentiated | High | >20 | >20% |
| NEC, large-cell type (LCNEC) | >20 | >20% | ||
| MiNEN | Well or poorly differentiated | Variable | Variable | Variable |
* mitotic rates are determined by counting in 50 fields of 0.2 mm2; ** Ki-67 values are expressed as the percentage of positive cells. Abbreviations: LCNEC, large-cell neuroendocrine carcinoma; MiNEN, mixed neuroendocrine–non-neuroendocrine neoplasm; NEC, neuroendocrine carcinoma; NET, neuroendocrine tumour; SCNEC, small-cell neuroendocrine carcinoma.
The hallmark of NENs is their expression of somatostatin receptors (SSTRs), as somatostatin inhibits cell growth and hormone secretion in normal and cancerous neuroendocrine cells [12]. Somatostatin receptors are G-protein-coupled receptors with a typical transmembrane domain that includes five distinct subtypes named 1 to 5, with the gene encoding for the SSTR2 also producing two splice variants, SSTR2 isoform A and B. While the natural ligands of SSTRs (i.e., somatostatin-14, somatostatin-28 and cortistatin) all bind to the receptors with high affinity, somatostatin analogues (SSAs)-octreotide, vapreotide and lanreotide-bind only to SSTR2 and with a lower affinity to SSTR3 and 5 [13][14][15][13,14,15]. Neuroendocrine neoplasms express all SSTRs at different concentrations, with SSTR2 being the predominant receptor found across NENs of different origins, followed by SSTR3 in gastroenteropancreatic (GEP)-NENs and SSTR1 and SSTR5 in midgut NENs [16][17][18][16,17,18].
The GEP tract is the most common site for NENs, with the small intestine (SI) and the pancreas being the most prevalent sites of origin for more advanced neoplasms. For these neoplasms, treatment strategies are based on information on SSTRs expression, tumour stage and grade (including differentiation) and the expression of neuroendocrine biomarkers [19].
The definitive diagnosis of an NEN is made by histopathological examination of tumour tissue, obtained either via a biopsy or following surgery. Morphologic imaging, however, is essential as a baseline evaluation for staging, in particular for identifying the presence of metastases, while functional imaging is important to assess the functional and metabolic status of the tumour. Combining morphological (e.g., computer tomography-CT) and functional imaging techniques is fundamental in the decision-making process of the therapeutic approach to patients with GEP-NENs [20]. Gallium68 (68Ga)-DOTA-peptide positron emission tomography (PET)/CT, i.e., 68Ga-DOTATATE or 68Ga-DOTATOC, remains the gold standard for assessing the eligibility and response to peptide receptor radionuclide therapy (PRRT), especially for well-differentiated grade 1 and grade 2 GEP-NETs [21][22][21,22]. However, NENs often show heterogeneous expression of SSTR, which could lead to inferior outcomes following targeted treatment and subsequently influence relapse and progression of the disease [21][22][23][24][25][21,22,23,24,25]. High-grade lesions and metastases can have a lower expression of SSTRs which may not be fully assessed on receptor-based imaging alone.
Spatial and temporal heterogeneity should be taken into account in the assessment of NENs, as it is not uncommon for GEP primary and metastatic sites to show intra-tumour and inter-tumour heterogeneity in their Ki-67 index, as well as in their SSTR expression and cell signalling pathways, leading to incomplete understanding of their tumour biology and behaviour [26][27][28][29][30][31][26,27,28,29,30,31].
2. Tumour Heterogeneity in GEP-NENs
Tumour heterogeneity is a common phenomenon in GEP-NENs (Figure 1) and has a negative impact on treatment success and prognosis as it produces cell clones that do not express treatment targets (i.e., SSTR, mammalian target of rapamycin–mTOR- signalling pathway, Ki-67) [32][33].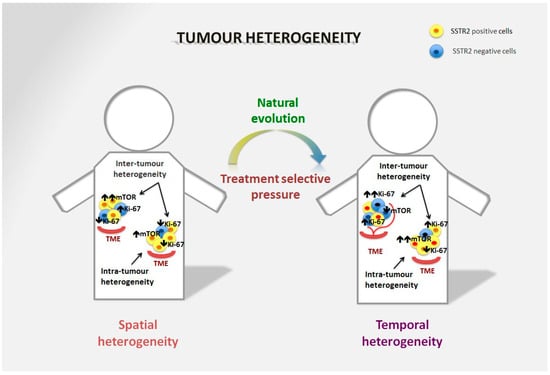
Figure 1. Spatial and temporal heterogeneity in NENs. Neuroendocrine neoplasms generally express SSTR2 on the tumour surface, and are well-differentiated tumours in the majority of cases. However, spatial heterogeneity within the primary tumour may lead to the presence of areas with lower expression of SSTR2 and/or a different Ki-67 index. This heterogeneity is also frequent in metastatic sites and can differ significantly from the primary lesion. The mTOR pathway is also commonly involved in the onset of the disease and is particularly relevant in distant metastases, although over time alternative pathways may be involved in tumour survival. Moreover, temporal heterogeneity that can be linked to treatment selective pressure may lead to significant changes in tumour biology that affect prognosis and survival.