Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Jianxun Song and Version 2 by Conner Chen.
COVID-19 has become an increasing global health issue. Adaptive immune cells, especially T cells, have been extensively investigated in regard to SARS-CoV-2 infection. However, human health and T cell responses are also impacted by many other pathogens and chronic diseases.
- SARS-CoV-2
- T cells
- coinfection
1. Introduction
Since its start at the end of 2019, the COVID-19 outbreak has posed significant health risks to the human population, surpassing more than 6.5 million deaths across the world (WHO cumulative data up to Oct 2022). Over the past 3 years, publications regarding coinfection with SARS-CoV-2 and other pathogens, such as viruses, bacteria, and parasites, have been reported. Tissue damage resulting from SARS-CoV-2 includes lung failure [1], brain damage [2], intestinal mucosal damage [3], and cardiovascular disease [4]. Furthermore, in cases when a cytokine storm is present in SARS-CoV-2 infection, coinfection may exacerbate the outcomes of other types of diseases, such as allergy, diabetes, and hypertension.
T cells play a pivotal role during viral infection and immune-related diseases. Due to SARS-CoV-2’s ability to alter the T cell response, it may affect the outcomes of several common pathogenic infections and chronic diseases. In a reported case of HIV coinfection with SARS-CoV-2, depletion of CD4+ T cells was observed to affect the patient’s clinical outcome [5]. In other conditions such as TB infection and diabetes, there is an increased risk of severe SARS-CoV-2 due to exacerbation of symptoms [6]. In diabetic patients, the increased clinical risk is positively correlated with SARS-CoV-2 infection [7].
2. SARS-CoV-2 Infection and T Cell Response
Within lethal contagious respiratory viruses, the most prevalent transmission route is an airborne infection [8]. In Figure 1, when the SARS-CoV-2 reaches the lung’s epithelial layer, the spike protein on the viral surface will bind with the ACE2 receptor, accelerating the entry of the virus [9][10][9,10]. Fusion of the virus and host cell membrane results in the release of the viral genome [11]. Essential components (including N, S, M, and E proteins) and sub-genomic viral RNA are subsequently generated and assembled to form an intact virus [12]. The newly generated virus is released from the cytoplasm through exocytosis [12] and SARS-CoV-2 will be captured by antigen-presenting cells (APC). Viral antigens are then processed and presented to the T cells [13][14][13,14]. Meanwhile, the defense mechanism mediated by T cells with SARS-CoV-2 is different from the antibody-dependent response due to their ability to recognize a broad range of viral epitopes [13][15][13,15]. Compared with the common cold, SARS-CoV-2 infection generates a diverse epitope pool and increases the frequency of both viral-epitope-specific CD4+ and CD8+ T cells among convalescent SARS-CoV-2 patients [13]. In terms of infection, T cells secrete IFNγ, Granzyme B, and TNFα to help eliminate SARS-CoV-2 infected cells [16]. In mild COVID-19 patients, the effector CD4+ or CD8+ T cells will proliferate and form a defense mechanism during the acute phase [17]. In moderate-symptom patients, the natural killer T cell (NKT) CD160 population responds quickly by direct cytotoxicity [18]. Furthermore, the SARS-CoV-2-specific memory T cells are beneficial for providing essential protection against future reinfection [15][19][20][21][15,19,20,21].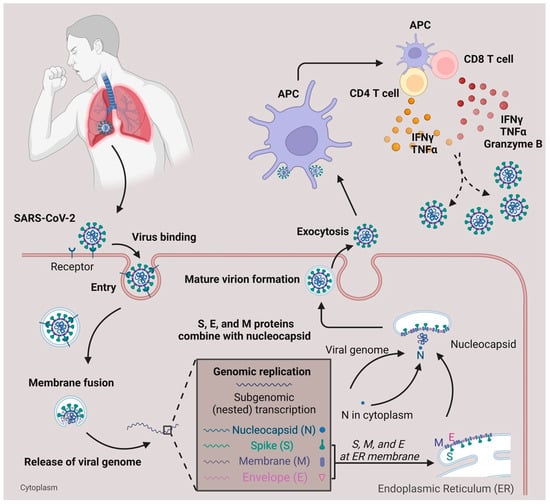
Figure 1. SARS-CoV-2 infection and T cell response. SARS-CoV-2 infects the respiratory tract by binding to the ACE2 surface receptor. Following entry via membrane fusion, it releases its viral genome in the cytoplasm. Using the host cell machinery, viral genome replication, and subgenomic transcription generate essential components needed for the virus to pack together. After the maturation of the virion, it is released from the host cell through exocytosis. When the viruses are captured and processed by antigen-presenting cells (APC), the T cells, either CD4+ or CD8+, are activated and secrete cytokines (IFNγ, TNFα, Granzyme B) to defend against viruses.
3. T Cell Subsets and Surface Markers Change during SARS-CoV-2 Infection
As studied, SARS-CoV-2 patients with severe symptoms often displayed progressive lymphopenia, while patients with mild symptoms showed normal absolute lymphocyte counts [22]. Within the CD8+ T cell populations in these patients, an increased population of CD8+ effector memory T cells was observed [22]. On the other hand, lower CD4+ or CD8+ frequencies were detected in severe COVID-19 patients compared to mild COVID-19 patients [23]. A study performed by Neidleman et al. indicated that CD27+CD28+CD8+ T effector memory cells re-expressing CD45RA (TEMRA) were predominant in the recovery period of SARS-CoV-2 patients, contradicting the expression of the CD27 marker [24]. Moreover, there was an increase in both central memory CD8+ T cells (CD45RA−CD27+CCR7+) and effector memory CD8+ T cells (CD45RA−CD27+CCR7−) [25][26][27][25,26,27]. In regards to Ki67+ and HLA-DR markers co-expressed on CD8+ T cells, SARS-CoV-2 infection led to an increase in Ki67+HLA-DR+CD8+ T cells in the patients [26]. In addition, there was an increase in the expression of activation markers, including HLA-DR and CD45RO, on CD8 T cells in severe SARS-CoV-2 patients compared to mild patients [28]. An increased proportion of CD38hi+ CD8+ T cells was also observed in severe SARS-CoV-2 infected patients [26][29][30][26,29,30]. In addition, a reduction in CD27+CD8+ T cells and an increase in CD127+CD8+ T cells were found in SARS-CoV-2 patients [24]. When compared to healthy blood donors, CTLA-4, LAG-3, and Tim-3 were significantly expressed in memory CD8+ T cells from patients with severe SARS-CoV-2 infection, while there was no difference for the inhibitory immune checkpoint PD-1 marker [26][30][26,30]. Cords detected that LAG-3 and TIGIT expression were upregulated in SARS-CoV-2-specific CD4+ T cells when compared to CD4+ T cells in healthy patients [31]. A higher percentage of cells expressing HLA-DR and CD45RO were also observed within the effector CD4+ T cell population in severe patients compared to mild patients [32]. On the other hand, a reduction in CD28+CD4+ T cells was detected in severe patients compared to mild patients [32]. Cytokines, including IL-2R, IL-6, IL-8, and IL-10, experienced an increase in production from effector CD4+ T cells in severe SARS-CoV-2 patients [33][34][33,34]. The decreased expression of CD45RA and CCR7 indicated that T central memory cells are more dominant than T effector memory cells [24]. It was also reported that there were high populations of memory CD4+ T cells expressing CD38, CD69, Ki-67, and PD-1 in memory CD4+ T cells from patients with severe SARS-CoV-2 infection compared to healthy blood donors [26]. Among CD4+ T cell subsets, CD4+ Teff cells differentiate into functionally diverse subsets such as Th1, Th2, Tfh, Th17, and Treg cells. T helper 1 (Th1) cells protect against intracellular parasites by the secretion of IFNγ [35][36][35,36]. Studies have indicated that Th1 cells predominate during SARS-CoV-2 infection through upregulation of IFNγ but not IL-4 or IL-17. T helper 2 (Th2) cells help to eliminate extracellular pathogens [36]. Studies also indicate that Th2 and Th17 cells play a role in SARS-CoV-2 infection in which Th2 cells were claimed to be unfavorable for recovery [24][30][37][24,30,37]. Lower levels of Th2 populations were observed in mild patients compared to healthy donors, but no difference was indicated between mild and severe patients. The disproportionate ratio of Th2/Th1 cells revealed a lead in severe infection, with abnormal secretion of associated cytokines such as IL-4, IL-5, IL-10, and IL-13 [30][38][39][40][30,38,39,40]. T follicular helper cells (Tfh), a particular subset of CD4+ T cells that regulates antibody responses with B cells, have been noted to play a role in SARS-CoV-2 infection [22]. Mathew et al. indicated that there was a decreased population of CD45RA−CXCR5+PD-1+ circulating Tfh (cTfh) cells post-SARS-CoV-2 infection with an upward trend of Tfh cell population in recovered patients within six months [27]. Among cTfh cells, there are multiple subsets including Tfh1, Tfh2, and Tfh17 that are categorized based on differences in the secretion of cytokines. A lower population of cTfh17 (CXCR5+CXCR3−CCR6+) cells, which secrete IL-21 and IL-17A, was discovered in a common variable immunodeficiency disease (CVID) patient with SARS-CoV-2 infection compared to the immunocompetent donor [22][41][22,41]. However, for PD-1 marker expression, there were multiple contradictory studies indicating that there were no changes in PD-1+ Tfh cells in SARS-CoV-2 patients [42]. A high expression of ICOS was also found in SARS-CoV-2-specific Tfh cells [24]. In regard to Treg cells, which act as suppressors in the immune system, there are conflicting studies. An increased percentage of regulatory T cells (CD3+CD4+CD25+CD127low) and a higher secretion of IL-10, TGF-b, IL-6, and IL-18 were detected during COVID-19 infection, specifically with N peptide stimulation [43][44][43,44]. Increased populations of Treg cells were present in mildly symptomatic patients compared to healthy controls [43].4. Role of T Cells during Coinfection with Viruses, Bacteria, and Parasites
During the three-year pandemic, researchers have noticed cases of coinfection with SARS-CoV-2 and other viruses (e.g., HBV, HIV, HCV, and influenza), bacteria (e.g., Mtb), and parasites (e.g., protozoa and helminths) (Figure 23).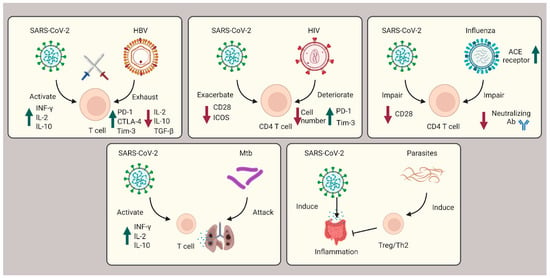
Figure 23. Role of T cells during coinfection with viruses, bacteria, and parasites. In SARS-CoV-2 coinfection with HBV, exhausted T cells are caused by HBV infection and subsequently countered with SARS-CoV-2 infection cytokine storms. In HIV coinfection, CD4+ T cells are corrupted, leading to exacerbated patient outcomes. Influenza coinfection decreases neutralization antibody efficacy and increases ACE2 receptors, overall boosting SARS-CoV-2 infection. For Mtb coinfection, both infections will affect the lung tissue, resulting in respiratory failure. In coinfection with parasites, Th2 and Treg cells are induced to suppress the immune system and ameliorate the intestinal inflammation severity of SARS-CoV-2.