Schistosomiasis, also known as bilharzia, is a major parasitic disease caused by blood flukes (trematode worms) of the genus Schistosoma that live in fresh waterways in tropical and subtropical zones. Over 200 million people are infected globally, 90% of whom live in sub-Saharan Africa. Nigeria has the highest burden of schistosomiasis in this region. Elimination programmes have focused on human infections, with limited attention on infections in livestock that could be transmissible to humans, i.e. zoonotic schistosomiasis. This is now recognized as a risk factor for increased transmission and recrudescence of infection of more than one schistosome species, as well as of potential hybrid variants. Members of farming communities who herd grazing cattle, goats and sheep in proximity to rivers containing Bulinus freshwater snails that are the intermediate host of asexual lifecycle stages are particularly at risk of becoming infected through daily contact with contaminated water.
- Schistosoma
- schistosome
- zoonosis
- zoonotic
- livestock
- parasite
- Nigeria
- West Africa
- snail
- transmission
1. Introduction
2. Life Cycle of Schistosoma haematobium
The life cycle of S. haematobium passes through two phases, asexual and sexual, which involve infection of the snail intermediate host and the vertebrate definitive host, respectively. Typically, the cycle starts when eggs released with urine from an infected host come into contact with a freshwater body harbouring snails belonging to the genus Bulinus. The eggs hatch immediately to release free-swimming miracidia that penetrate the host. Upon successful establishment, these develop asexually through several stages into cercariae that leave the snail in search of the definitive host. While having a very short life span, a single miracidium established in a suitable snail host produces about 200 cercariae daily [19,20][19][20]. Cercariae penetrate the skin of suitable definitive hosts who come into contact with the contaminated water body, whereupon they transform into schistosomula. Usually, this penetration takes place in surface water, where people and livestock congregate daily for domestic and recreational purposes [21]. The schistosomula migrate through the epidermis and dermis, entering the blood to home to the lungs [20,22,23,24][20][22][23][24]. Subsequently, schistosomula exit the lungs to the left side of the heart through the pulmonary veins and enter the abdominal aorta, where they either pass through the coeliac trunk, the inferior and superior mesenteric arteries, or the iliac arteries to reach the portal veins of the liver [24]. Here, schistosomula lose their migratory ability, grow, and develop into adult male and female pairs [24]. Thereafter, a pair travels against the blood flow in the venous circulation, settles in the vesical venous plexus and produces eggs that migrate to the bladder, ureter, and other parts of the urinary tract [20,24][20][24]. These eggs permeate the walls of blood vessels, bladder, or genital organs [24]. Those that access the bladder are passed out with urine into a body of freshwater, where hatched miracidia continue the transmission cycle [20]. Other Schistosoma species follow a similar life cycle, but with different predilection sites and egg morphologies [25,26][25][26].3. Zoonotic Schistosomiasis
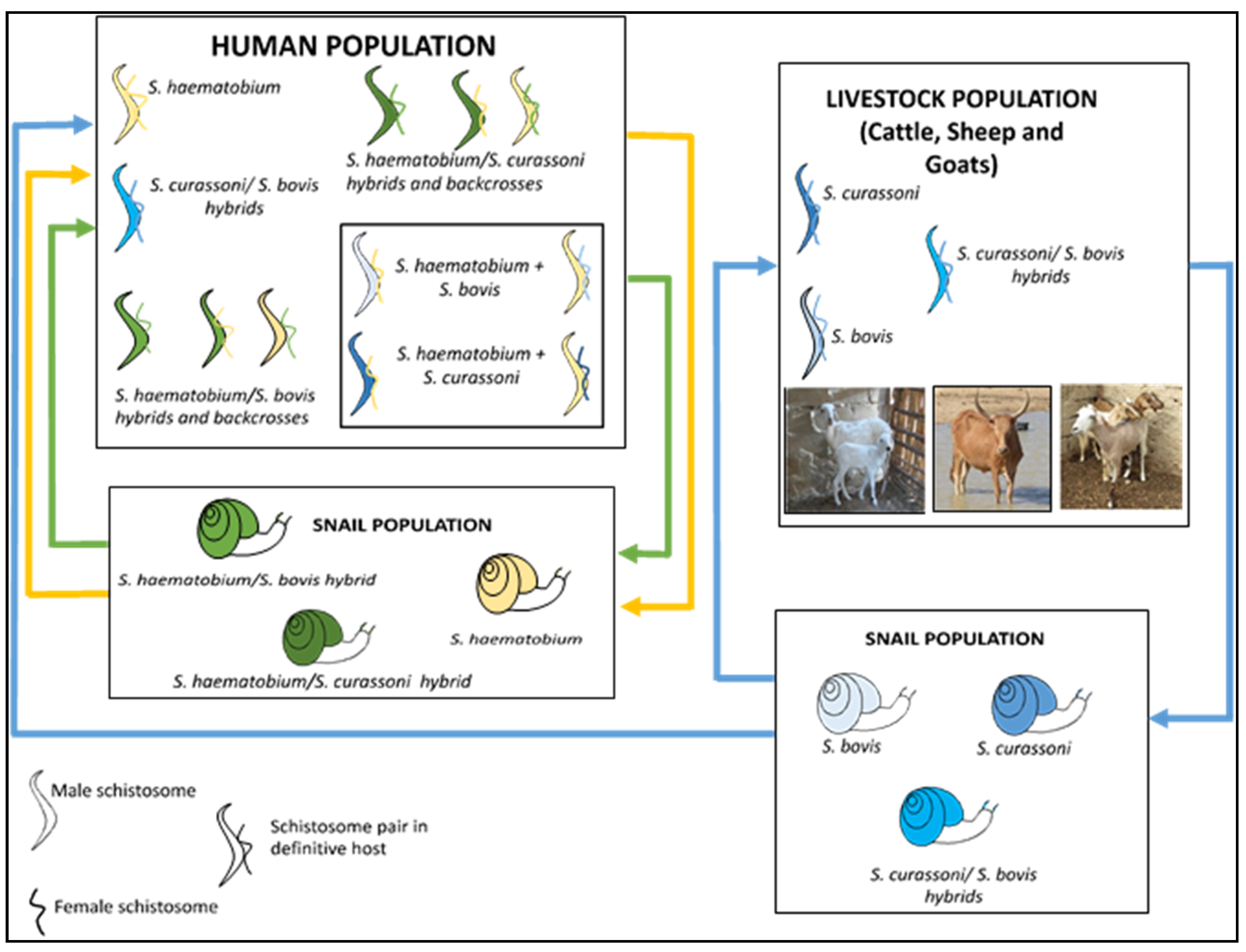
The terms zoonosis or zoonotic disease are used to classify a range of infectious diseases and their causative agents that are capable of being transmitted and subsequently established between human and animal hosts [27]. Hence, zoonotic schistosomiasis refers to the disease condition in which a Schistosoma species known to affect a particular human or animal host is found in another host not specific to the schistosome group [13,28][13][28]. These species are naturally shared between humans and other animal hosts as a result of extensive mixed interactions at transmission sites [29,30][29][30] (Figure 1).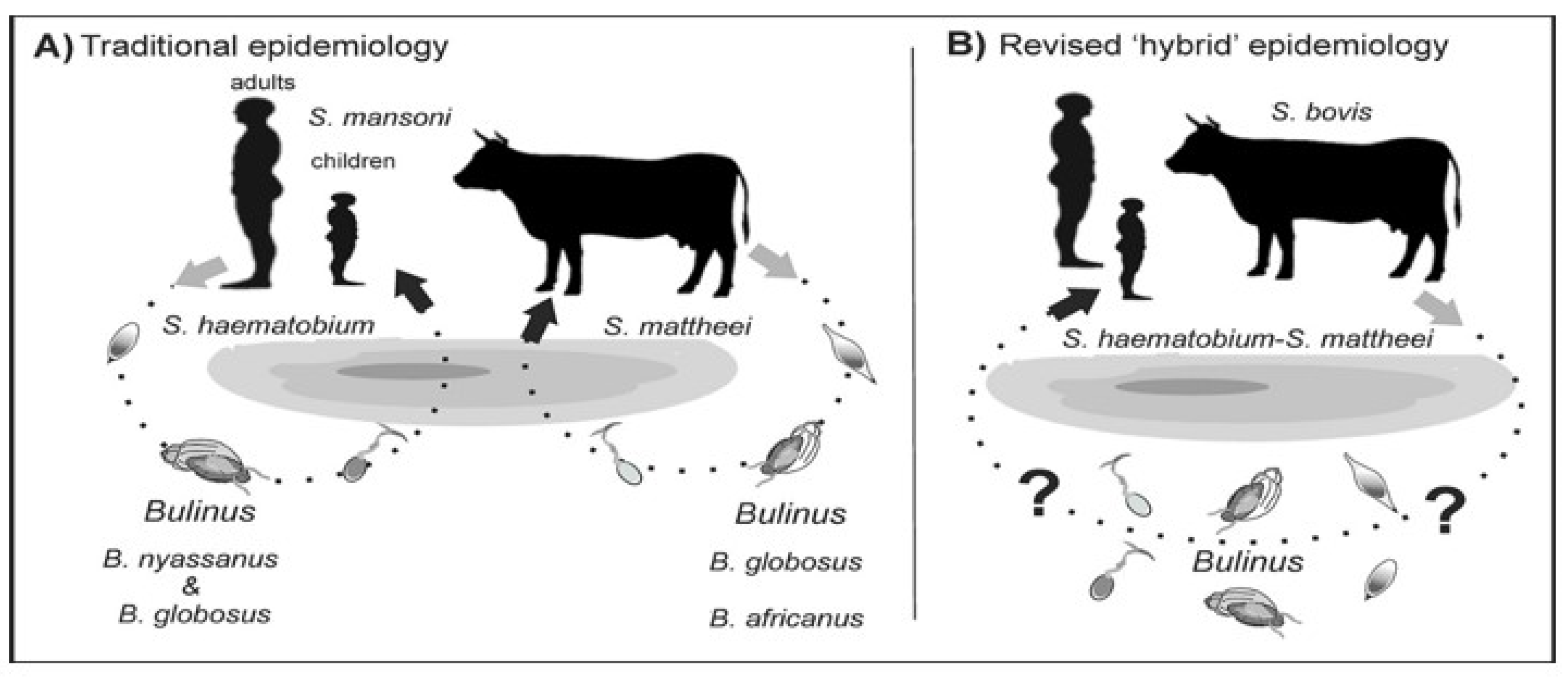
4. Schistosomiasis Control Programme in Nigeria
The National Control Schistosomiasis Programme was established by the Federal Government of Nigeria in 1988, following which epidemiological surveying revealed that an estimated 20 million people were infected. Five pilot control projects were initiated in the states of Borno, Ebonyi, Katsina, Kwara, and Ondo, which have varying ecological predispositions to schistosomiasis. However, due to limited funds, the successes of these pilot projects have fallen short of initially planned targets. In 2020, the Carter Center incorporated schistosomiasis control activities into an ongoing onchocerciasis eradication programme in Plateau and Nasarawa states [35]. A decade before this, mass administration of praziquantel (MDA) commenced in other states with the support of the WHO, UNICEF, and other non-governmental development organizations. To date, planning and implementation of MDA occur within implementation units (IUs), which may be a district, province, or local government area [9]. In common with policy for combatting soil-transmitted helminth infections [36], disease prevalence and other population data generated at the community level are usually aggregated at IUs. For schistosomiasis, this information is used to determine praziquantel thresholds to be implemented [37]. MDA takes place biannually for all school-age children in districts where schistosomiasis prevalence is above 50%. MDA occurs annually when the prevalence is between 10 and 49.9%, and biennially when the prevalence is between 1 and 10% [9]. However, recent WHO guidelines on schistosomiasis risk classification have revised these recommendations as follows. In endemic communities with the prevalence of Schistosoma spp. infection ≥ 10%, in order to control schistosomiasis morbidity and advance towards eliminating the disease as a public health problem, the WHO recommends annual preventive chemotherapy with a single dose of praziquantel at ≥75% treatment coverage in all age groups from 2 years old, including adults, pregnant women after the first trimester, and lactating women [9]. In endemic communities with a prevalence of Schistosoma spp. infection ≥ 10% that demonstrate a lack of an appropriate response to annual preventive chemotherapy, despite adequate treatment coverage (≥75%), the WHO suggests consideration of preventive chemotherapy biannually (twice a year) instead of annually [38]. The review of schistosomiasis treatment data over recent years (2014–2021) shows that the number of districts treated is significantly lower than those requiring treatment. Treatment coverage was significantly lower in 2021 (during the height of the COVID-19 pandemic) than experienced in the previous 7 years. In addition, less than half of the districts met the 75% effective coverage threshold in the last 3 successive years (2019–2021). By endemicity, the proportion of districts with high prevalence (>50%) reduced from (n = 15, 3%) in 2014 to (n = 10, 2%) in 2021, and those with moderate prevalence (10–49%) reduced from (n = 305, 52%) in 2014 to (n = 294, 50%) in 2021. However, the proportion of districts with low prevalence (<10%) increased from (n = 263, 48%) in 2014 to (n = 279, 48%) in 2021 [39]. The poor coverage of praziquantel across the years is most likely connected to the availability of medicines and logistics required to drive mass treatment campaigns [40]. This is particularly worsened by the myriad of issues surrounding the availability, motivation and retention of drug distributors/health workers who are major stakeholders in the campaign [41]. These observations are in line with a 2021 global report on schistosomiasis treatment, as only 30% of people (i.e., 75.3 million of the 251.4 million Nigerians requiring praziquantel) were treated [40]. The ongoing pandemic and efforts targeted at mitigating its impacts have decreased the provision of NTD interventions, particularly praziquantel medicines for schistosomiasis. Nevertheless, there is a need for more concerted efforts towards increasing coverage of medicines in line with achieving the 2030 global targets of the WHO NTD elimination roadmap. These include: (1) reducing the proportion of moderate and heavy intensity infections to <1% in 78 countries; (2) reducing by 50% the number of tablets required during MDA; and (3) increasing domestic financial support for MDA [3,4][3][4]. Meeting these targets is largely dependent on increased community participation during MDA, with absolute geographical reach and high programme coverage [42,43,44,45][42][43][44][45].5. Implication of Hybridization for Schistosomiasis Control Efforts
The existence of hybrid schistosomes is of great concern to the schistosomiasis control programme in Nigeria, with possible implications for widening host range, increased transmission potential, altered pathology, and drug resistance [46]. Interactions between different Schistosoma species are known to improve reproductive capacity, as manifested by an increased number of parasite offspring, a faster maturation time, and a larger intermediate host range [47,48][47][48]. A major concern is that with the emergence of hybrid schistosomes, the efficacy of praziquantel treatment might be lowered [49]. This has been reported under both field and laboratory conditions for S. mansoni [50,51,52][50][51][52] and was linked to the cryptic role played by hybridization. The success achieved by using this drug is therefore threatened by the outbreak of Schistosoma hybrids [5]. Furthermore, genetic diversity among the progeny of zoonotic schistosomes offers better phenotypic characteristics compared to those of either parent [53], thereby enhancing the exploitation of the host. Certain genotypic traits of the zoonotic hybrid parasite show refined adaptations to better avoid recognition and to resist the host’s adaptive immune system, potentially giving rise to greater infectivity and unusual pathologies [54]. Infection of multiple animal reservoir hosts by zoonotic hybrid schistosomes makes the elimination of the disease more challenging [15,16][15][16]. The difficulty to control and prevent Schistosoma hybrid infections in some endemic areas may be due to the ubiquitous presence of rodents, which serve as the reservoir host for many schistosome species, thereby readily enabling co-infection [55]. The magnitude of the contribution of non-human mammalian hosts to the transmission of schistosomiasis remains understudied, especially for species other than S. japonicum. This has been highlighted by the WHO, which recommends testing for Schistosoma infections in non-human mammalian hosts [38].References
- Hotez, P.J.; Fenwick, A. Schistosomiasis in Africa: An emerging tragedy in our new global health decade. PLoS Negl. Trop Dis. 2009, 3, e485.
- Hotez, P.J.; Kamath, A. Neglected tropical diseases in sub-Saharan Africa: Review of their prevalence, distribution, and disease burden. PLoS Negl. Trop. Dis. 2009, 3, e412.
- World Health Organization. Schistosomiasis. 8 January 2022. Available online: https://www.who.int/news-room/fact-sheets/detail/schistosomiasis (accessed on 30 January 2023).
- World Health Organization. Ending the Neglect to Attain the Sustainable Development Goals: A Road Map for Neglected Tropical Diseases 2021–2030; World Health Organization: Geneva, Switzerland, 2020.
- Stothard, J.R.; Kayuni, S.A.; Al-Harbi, M.H.; Musaya, J.; Webster, B.L. Future schistosome hybridizations: Will all Schistosoma haematobium hybrids please stand-up! PLoS Negl. Trop. Dis. 2020, 14, e0008201.
- Engels, D.; Chitsulo, L.; Montresor, A.; Savioli, L. The global epidemiological situation of schistosomiasis and new approaches to control and research. Acta Trop. 2002, 82, 139–146.
- Centers for Disease Control and Prevention. DPDx—Laboratory Identification of Parasites of Public Health Concern. Schistosomiasis. Available online: https://www.cdc.gov/dpdx/schistosomiasis/index.html (accessed on 30 January 2023).
- Ekpo, U.F.; Hürlimann, E.; Schur, N.; Oluwole, A.S.; Abe, E.M.; Mafe, M.A.; Nebe, O.J.; Isiyaku, S.; Olamiju, F.; Kadiri, M.; et al. Mapping and prediction of schistosomiasis in Nigeria using compiled survey data and Bayesian geospatial modelling. Geospat. Health 2013, 7, 355–366.
- World Health Organization. Helminth Control in School-Age Children: A Guide for Managers of Control Programmes, 2nd ed.; World Health Organization: Geneva, Switzerland, 2011; Available online: https://apps.who.int/iris/bitstream/handle/10665/44671/9789241548267_eng.pdf (accessed on 30 January 2023).
- Rollinson, D. A wake-up call for urinary schistosomiasis: Reconciling research effort with public health importance. Parasitology 2009, 136, 1593–1610.
- Webster, B.L.; Southgate, V.R. Mating interactions of Schistosoma haematobium and S. intercalatum with their hybrid offspring. Parasitology 2003, 126, 327–338.
- Léger, E.; Garba, A.; Hamidou, A.A.; Webster, B.L.; Pennance, T.; Rollinson, D.; Webster, J.P. Introgressed animal schistosomes Schistosoma curassoni and S. bovis naturally infecting humans. Emerg. Infect. Dis. 2016, 22, 2212–2214.
- Huyse, T.; Webster, B.L.; Geldof, S.; Stothard, J.R.; Diaw, O.T.; Polman, K.; Rollinson, D. Bidirectional introgressive hybridisation between cattle and human schistosome species. PLoS Pathog. 2009, 5, e1000571.
- Soentjens, P.; Cnops, L.; Huyse, T.; Yansouni, C.; Vos, D.D.; Bottieau, E.; Clerinx, J.; Esbroeck, M.V. Diagnosis and clinical management of Schistosoma haematobium–Schistosoma bovis hybrid infection in a cluster of travelers returning from Mali. Clin. Infect. Dis. 2016, 63, 1626–1629.
- Savassi, B.A.E.S.; Mouahid, G.; Lasica, C.; Mahaman, S.-D.K.; Garcia, A.; Courtin, D.; Allienne, J.-F.; Ibikounlé, M.; Moné, H. Cattle as natural host for Schistosoma haematobium (Bilharz, 1852) Weinland, 1858 x Schistosoma bovis Sonsino, 1876 interactions, with new cercarial emergence and genetic patterns. Parasitol. Res. 2020, 119, 2189–2205.
- Savassi, B.A.E.S.; Dobigny, G.; Etougbétché, J.R.; Avocegan, T.T.; Quinsou, F.T.; Gauthier, P.; Ibikounlé, M.; Moné, H.; Mouahid, G. Mastomys natalensis (Smith, 1834) as a natural host for Schistosoma haematobium (Bilharz, 1852) Weinland, 1858 x Schistosoma bovis Sonsino, 1876 introgressive hybrids. Parasitol. Res. 2021, 120, 1755–1770.
- Tchuenté, L.; Southgate, V.; Njiokou, F.; Njiné, T.; Kouemeni, L.; Jourdane, J. The evolution of schistosomiasis at Loum, Cameroon: Replacement of Schistosoma intercalatum by S. haematobium through introgressive hybridisation. Trans. R. Soc. Trop. Med. Hyg. 1997, 91, 664–665.
- Webster, B.L.; Tchuenté, L.T.; Jourdane, J.; Southgate, V.R. The interaction of Schistosoma haematobium and S. guineensis in Cameroon. J. Helminthol. 2005, 79, 193–197.
- Grimes, J.E.; Croll, D.; Harrison, W.E.; Utzinger, J.; Freeman, M.C.; Templeton, M.R. The roles of water, sanitation and hygiene in reducing schistosomiasis: A review. Parasit Vectors 2015, 8, 156.
- Nelwan, M.L. Schistosomiasis: Life cycle, diagnosis, and control. Curr. Ther. Res. Clin. Exp. 2019, 91, 5–9.
- Mafiana, C.F.; Ekpo, U.F.; Ojo, D.A. Urinary schistosomiasis in preschool children in settlements around Oyan Reservoir in Ogun State, Nigeria: Implications for control. Trop. Med. Int. Health 2003, 8, 78–82.
- Curwen, R.S.; Wilson, R.A. Invasion of skin by schistosome cercariae: Some neglected facts. Trends Parasitol. 2003, 19, 63–66.
- He, Y.X.; Salafsky, B.; Ramaswany, K. Comparison of skin invasion among three major species of Schistosoma. Trends Parasitol. 2005, 21, 201–203.
- Nation, C.S.; Da’dara, A.A.; Marchant, J.K.; Skelly, P.J. Schistosome migration in the definitive host. PLoS Negl. Trop. Dis. 2020, 14, e0007951.
- McManus, D.P.; Dunne, D.W.; Sacko, M.; Utzinger, J.; Vennervald, B.J.; Zhou, X.-N. Schistosomiasis. Nat. Rev. Dis. Prim. 2018, 4, 13.
- Centers for Disease Control and Prevention. Parasites—Schistosomiasis. In Biology; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2019. Available online: https://www.cdc.gov/parasites/schistosomiasis/biology.html (accessed on 30 January 2023).
- Chomel, B.B. Control and prevention of emerging parasitic zoonoses. Int. J. Parasitol. 2008, 38, 1211–1217.
- Huyse, T.; Van Den Broeck, F.; Hellemans, B.; Volckaert, F.A.M.; Polman, K. Hybridisation between the two major African schistosome species of humans. Int. J. Parasitol. 2013, 43, 687–689.
- Standley, C.J.; Mugisha, L.; Dobson, A.P.; Stothard, J.R. Zoonotic schistosomiasis in non-human primates: Past, present and future activities at the human–wildlife interface in Africa. J. Helminthol. 2012, 86, 131–140.
- Léger, E.; Webster, J.P. Hybridizations within the Genus Schistosoma: Implications for evolution, epidemiology and control. Parasitology 2017, 144, 65–80.
- Fan, P.C.; Lin, L.H. Hybridization of Schistosoma mansoni and Schistosoma japonicum in mice. Southeast Asian J. Trop. Med. Public Health 2005, 36, 89–96.
- Boissier, J.; Moné, H.; Mitta, G.; Bargues, M.D.; Molyneux, D.; Mas-Coma, S. Schistosomiasis reaches Europe. Lancet Infect. Dis. 2015, 15, 757–758.
- Steinauer, M.L.; Hanelt, B.; Mwangi, I.N.; Maina, G.M.; Agola, L.E.; Kinuthia, J.M.; Mutuku, M.W.; Mungai, B.N.; Wilson, W.D.; Mkoji, G.M.; et al. Introgressive hybridization of human and rodent schistosome parasites in western Kenya. Mol. Ecol. 2008, 17, 5062–5074.
- Borlase, A.; Webster, J.P.; Rudge, J.W. Opportunities and challenges for modelling epidemiological and evolutionary dynamics in a multihost, multiparasite system: Zoonotic hybrid schistosomiasis in West Africa. Evol. Appl. 2017, 11, 501–515.
- The Carter Center. Schistosomiasis Control Program. 2021. Available online: https://www.cartercenter.org/resources/pdfs/factsheets/schistosomiasis-facts.pdf (accessed on 30 January 2023).
- Mogaji, H.O.; Dedeke, G.A.; Bada, B.S.; Bankole, S.; Adeniji, A.; Fagbenro, M.T.; Omitola, O.O.; Oluwole, A.S.; Odoemene, N.S.; Abe, E.M.; et al. Distribution of ascariasis, trichuriasis and hookworm infections in Ogun State, Southwestern Nigeria. PLoS ONE 2020, 15, e0233423.
- World Health Organization Regional Office for Africa. Expanded Special Project for Elimination of Neglected Tropical Diseases (ESPEN). Optimizing Schistosomiasis MDA Implementation in Countries. Data Analysis Tool, June–July 2019. Available online: https://espen.afro.who.int/system/files/content/resources/Schistosomiasis%20Data%20analysis%20tool%20-%20Presentation%20%2820190724_English%29.pdf (accessed on 30 January 2023).
- WHO. Guideline on Control and Elimination of Human Schistosomiasis; World Health Organization: Geneva, Switzerland, 2022; Available online: https://apps.who.int/iris/rest/bitstreams/1410449/retrieve (accessed on 30 January 2023).
- World Health Organization; Regional Office for Africa. Expanded Special Project for Elimination of Neglected Tropical Diseases (ESPEN). Nigeria Overview. Available online: https://espen.afro.who.int/countries/nigeria (accessed on 30 January 2023).
- World Health Organization. Schistosomiasis and soil-transmitted helminthiases: Progress report, 2021. Wkly. Epidemiol. Rec. 2022, 97, 621–632. Available online: https://www.who.int/publications/i/item/who-wer9748-621-632 (accessed on 30 January 2023).
- Krentel, A.; Gyapong, M.; Mallya, S.; Boadu, N.Y.; Amuyunzu-Nyamongo, M.; Stephens, M.; McFarland, D.A. Review of the factors influencing the motivation of community drug distributors towards the control and elimination of neglected tropical diseases (NTDs). PLoS Negl. Trop. Dis. 2017, 11, e0006065.
- El-Setouhy, M.; Abd Elaziz, K.M.; Helmy, H.; Farid, H.A.; Kamal, H.A.; Ramzy, R.M.R.; Shannon, W.D.; Weil, G.J. The effect of compliance on the impact of mass drug administration for elimination of lymphatic filariasis in Egypt. Am. J. Trop. Med. Hyg. 2007, 77, 1069–1073.
- Olamiju, O.J.; Olamiju, F.O.; Adeniran, A.A.; Mba, I.C.; Ukwunna, C.C.; Okoronkwo, C.; Ekpo, U.F. Public awareness and knowledge of neglected tropical diseases (NTDs) control activities in Abuja, Nigeria. PLoS Negl. Trop. Dis. 2014, 8, e3209.
- Assaré, R.K.; N’Tamon, R.N.; Bellai, L.G.; Koffi, J.A.; Mathieu, T.-B.I.; Ouattara, M.; Hürlimann, E.; Coulibaly, J.T.; Diabaté, S.; N’Goran, E.K.; et al. Characteristics of persistent hotspots of Schistosoma mansoni in western Côte d’Ivoire. Parasit Vectors 2020, 13, 337.
- Mogaji, H.O.; Odoh, I.M.; Iyeh, C.I.; Adeniran, A.A.; Oyedeji, S.I.; Okoh, H.I.; Bayegun, A.A.; Omitola, O.O.; Umunnakwe, C.U.; Olamiju, F.O.; et al. Attendee’s awareness about preventive chemotherapy neglected tropical diseases (PC-NTD) control during the first world neglected tropical diseases day in Ekiti State, Nigeria. PLoS Negl. Trop. Dis. 2021, 15, e0009315.
- King, K.C.; Stelkens, R.B.; Webster, J.P.; Smith, D.F.; Brockhurst, M.A. Hybridization in parasites: Consequences for adaptive evolution, pathogenesis, and public health in a changing world. PLoS Pathog. 2015, 11, e1005098.
- Moné, H.; Minguez, S.; Ibikounlé, M.; Allienne, J.-F.; Massougbodji, A.; Mouahid, G. Natural interactions between S. haematobium and S. guineensis in the Republic of Benin. Sci. World J. 2012, 2012, 793420.
- Webster, B.L.; Diaw, O.T.; Seye, M.M.; Webster, J.P.; Rollinson, D. Introgressive hybridization of Schistosoma haematobium group species in Senegal: Species barrier break down between ruminant and human schistosomes. PLoS Negl. Trop. Dis. 2013, 7, e2110.
- Webster, J.P.; Molyneux, D.H.; Hotez, P.J.; Fenwick, A. The contribution of mass drug administration to global health: Past, present and future. Philos. Trans./R Soc. Lond. B—Biol. Sci. 2014, 369, 20130434.
- Lamberton, P.H.L.; Hogan, S.C.; Kabatereine, N.B.; Fenwick, A.; Webster, J.P. In vitro praziquantel test capable of detecting reduced in vivo efficacy in Schistosoma mansoni human infections. Am. J. Trop. Med. Hyg. 2010, 83, 1340–1347.
- Valentim, C.L.L.; Cioli, D.; Chevalier, F.D.; Cao, X.; Taylor, A.B.; Holloway, S.P.; Pica-Mattoccia, L.; Guidi, A.; Basso, A.; Tsai, I.J.; et al. Genetic and molecular basis of drug resistance and species-specific drug action in schistosome parasites. Science 2013, 342, 1385–1389.
- Webster, B.L.; Diaw, O.T.; Seye, M.M.; Faye, D.S.; Stothard, J.R.; Sousa-Figueiredo, J.C.; Rollinson, D. Praziquantel treatment of school children from single and mixed infection foci of intestinal and urogenital schistosomiasis along the Senegal River Basin: Monitoring treatment success and re-infection patterns. Acta Trop. 2013, 128, 292–302.
- Grigg, M.E.; Bonnefoy, S.; Hehl, A.B.; Suzuki, Y.; Boothroyd, J.C. Success and virulence in Toxoplasma as the result of sexual recombination between two distinct ancestries. Science 2001, 294, 161–165.
- Schelkle, B.; Faria, P.J.; Johnson, M.B.; van Oosterhout, C.; Cable, J. Mixed infections and hybridisation in monogenean parasites. PLoS ONE 2012, 7, e39506.
- Hanelt, B.; Mwangi, I.N.; Kinuthia, J.M.; Maina, G.M.; Agola, L.E.; Mutuku, M.W.; Steinauer, M.L.; Agwanda, B.R.; Kigo, L.; Mungai, B.N.; et al. Schistosomes of small mammals from the Lake Victoria Basin, Kenya: New species, familiar species, and implications for schistosomiasis control. Parasitology 2010, 137, 1109–1118.