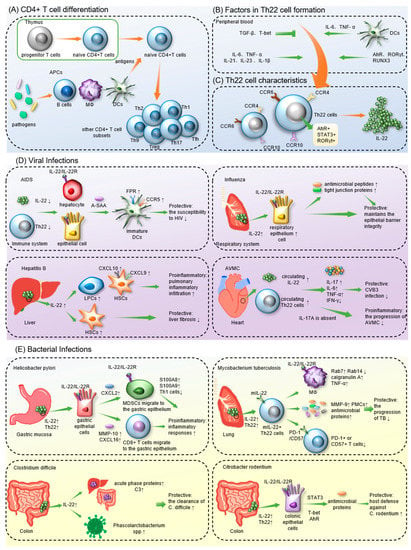
The differentiation process of Th22 cells and the production of IL-22 have been confirmed to be regulated by multiple factors, including cells, cytokines and transcription factors (Figure 1B).
According to previous studies
[36], the effects of IL-22 on host immune defense can be summarized in the following three aspects. (1) Mediating innate immunity. During
C. rodentium infection, IL-22 induces colonic epithelial cells to release the antibacterial proteins RegIIIβ and RegIIIγ
[37]. Other antimicrobial proteins, such as β-defensin 2, psoriasin (S100A7) and calgranulin A (S100A8), have also been shown to be upregulated by IL-22
[34][38][34,38]. IL-22 can mediate innate immunity by upregulating cell fluidity
[38][39][38,39], mucus secretion and the release of other cytokines
[11][40][11,40]. (2) Protecting the epithelial barrier and regulating cell proliferation. IL-22 can promote the production of tight junction proteins between epithelial cells to enhance the mucosal barrier
[41]. IL-22-induced STAT3 activation also maintains intestinal homeostasis by regulating wound healing of the intestinal mucosa
[42]. Additionally, when IL-17A is absent, IL-22 can inhibit epithelial cell apoptosis and promote epithelial proliferation and regeneration, exerting tissue-protective effects
[43][44][43,44]. IL-22 also activates the PI3K-Akt-mTOR pathway to inhibit pulmonary apoptosis and modulate the proliferation of normal human epidermal keratinocytes (NHEKs) and fibroblast-like synoviocytes (FLSs)
[45][46][45,46]. In addition, IL-22 can induce liver regeneration through suppressor cytokine signaling 3 (SOCS3), a STAT3-targeted gene
[47][48][47,48]. (3) Disrupting immune defenses through proinflammatory effects. During respiratory inflammation caused by Gram-negative bacteria
[44] and bleomycin
[43], IL-22 synergizes with IL-17A to recruit immune cells, resulting in extensive local inflammation. Moreover, proinflammatory acute-phase proteins can be released in human hepatoma cells after the injection of IL-22
[49]. However, in diseases such as AIDS, the acute phase proteins mediated by IL-22 are protective
[50]. Therefore, the ultimate effects of IL-22 are influenced by multiple factors, including the type of pathogen infected. Notably, IL-22 binding protein (IL-22BP), a natural IL-22 antagonist, can bind to IL-22 and block IL-22R recognition
[51][52][53][51,52,53]. Previous studies have revealed that additional IL-22BP neutralizes the effects of IL-22 and reverses the prognosis of diseases, making IL-22/IL-22BP a promising target for disease intervention
[51][54][55][51,54,55].
2. Th22 Cells in Infectious Diseases
2.1. Th22 Cells in Viral Infections
2.1.1. COVID-19
COVID-19 is pneumonia induced by severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). Studies in the context of infectious diseases have focused on the pathogenic mechanisms of COVID-19 since its outbreak in 2019. Hoffmann et al. proposed that COVID-19 shares some similar symptoms with influenza and respiratory syncytial virus (RSV)-induced pneumonia
[56]. According to previous studies, IL-22/Th22 is protective against influenza and RSV pneumonia
[54][57][58][54,57,58] and may exert a similar effect against COVID-19. Among patients with fulminant COVID-19-related myocarditis, some met the criteria for multisystem inflammatory syndrome (MIS-A
+), whereas the rest did not (MIS-A
-)
[59]. Compared to the MIS-A
- group, MIS-A
+ patients showed higher expression of IL-22, a better prognosis and lower mortality
[59]. These results suggest that IL-22 may have a protective and antiviral effect in MIS-A
+ COVID-19 patients. A novel study noted that abnormal dynamic IL-22R1 expression on blood myeloid cells and CD4+ T cells is a characteristic of SARS-CoV-2 infection
[60]. IL-22R1 expression on myeloid cells is discriminative for the severity of COVID-19
[60]. However, COVID-19 patients with different prognoses have similar IL-22 levels, suggesting that IL-22 does not affect the outcomes of SARS-CoV-2 infection
[61]. Furthermore, the number of IL-22R1-expressing myeloid cells is correlated with the plasma levels of COVID-19-related immune mediators
[60]. During the acute phase of COVID-19, the immune response leads to a dramatic increase in several cytokines, including IL-22, which is mainly produced by Th22 cells
[62][63][62,63]. This process is called cytokine release syndrome (CRS) and contributes to fatal complications, such as multiple organ failure and acute respiratory distress syndrome
[64][65][64,65]. This result indicated that IL-22R1
+ myeloid cells may participate in the cascade, leading to CRS and promoting the deterioration of COVID-19. This study also suggested that the IL-22-induced signaling pathway switches from protective to pathogenic as the disease progresses
[60]. Therefore, IL-22/Th22 cells may play a critical role in the pathological process of COVID-19, but the detailed mechanism still awaits further research.
2.1.2. AIDS
Acquired immunodeficiency syndrome (AIDS), an infectious illness that arises from infection with human immunodeficiency virus (HIV), is known for its high mortality and prolonged course
[66]. In comparison with healthy controls and HIV-infected patients, more acute-phase serum amyloid A (A-SAA) and IL-22 are produced in HIV-exposed but uninfected individuals (EUs)
[67][68][67,68]. IL-22 has been confirmed to promote the expression of A-SAA in epithelial cells and liver cells
[47][49][69][47,49,69]. Moreover, Misse’ et al. coincubated A-SAA with immature DCs in vitro for further exploration. A-SAA is an agonist of the formyl peptide receptor (FPR) and enhances FPR expression on DCs
[67]. The FPR promoted the phosphorylation of CCR5 and decreased the expression of CCR5 on DCs, resulting in decreased susceptibility of DCs to HIV
[67]. Therefore, it was suggested that the high resistance of EUs to HIV may be associated with IL-22-induced A-SAA.
However, the higher sensitivity of Th22 cells to HIV compared to other CD4+ T cell subsets is attributed to the high expression of CCR5 and α4β7 on Th22 cells
[70]. Both CCR5 and α4β7 are the main binding sites for HIV. Therefore, the counts and functional scores of Th22 cells are decreased significantly in the sigmoid colon of HIV patients
[71][72][71,72]. Moreover, since Th22/IL-22 limits the translocation of commensal bacteria into the systemic circulation
[73], Th22 depletion in HIV patients not only causes impairment of the intestinal barrier but also moves intestinal bacteria to the lamina propria
[71][72][71,72]. The translocated intestinal bacteria eventually enter the circulation and cause systemic immune activation, which is key to the pathogenesis of HIV
[72]. Furthermore, viral load in HIV-infected individuals is negatively associated with the level of serum IL-22, and one study indicated that IL-22 may inhibit HIV replication by regulating C-reactive protein (CRP) and IL-10
[74]. Compared to HIV patients with inflammatory disease coinfection, patients without inflammatory diseases showed lower neutrophil activation and a surplus of IL-22 expression
[75]. This result suggests that IL-22 controls HIV-related inflammatory injury by regulating neutrophil hyperactivation
[75]. In patients infected with HIV-2, a spontaneously attenuating HIV strain, the expression of CCL20 and CCL28 was significantly increased in the sigmoid colon, leading to local recruitment of Th22 cells
[76]. Therefore, Th22 cells counteract intestinal CD4+ T-cell exhaustion and maintain intestinal mucosal integrity. This finding may explain why HIV-2 infection is less dangerous than HIV-1 infection. In addition, mucosa-associated constant T-cell (MAIT) levels were positively correlated with Th22 cell frequency in HIV-infected children
[77]. Since innate immune responses are effectively regulated by MAITs
[78][79][78,79], Th22 cells may also be antiviral in HIV-infected children by mediating the level of MAITs. Moreover, compared to HIV-infected individuals with immune responses (IRs), patients without immune responses (INRs) expressed more IL-22 in the colon and showed more severe mucosal damage
[80]. After receiving antiretroviral therapy (ART), INRs had poorer immune recovery than IRs
[80], indicating that the dysregulation of IL-22 frequency may be responsible for the poor prognosis of INRs. Hence, although Th22 cells are downregulated in HIV patients, they modulate mucosal immunity and acute phase protein expression to suppress the progression of AIDS.
2.1.3. Hepatitis
Viral hepatitis is typically caused by infection with hepatitis B or C viruses
[81][82][81,82]. Both hepatitis B and C can increase the risk of cirrhosis and hepatocellular carcinoma
[81][83][81,83]. According to previous studies, IL-22 showed weak anti-HBV effects and no direct antiviral activity against hepatitis C virus (HCV)
[84]. Surprisingly, IL-22
-/- mice showed increased susceptibility to inflammatory liver damage, demonstrating that IL-22 protects hepatocytes through mechanisms other than direct virus killing
[85].
In HBV-infected patients, both intrahepatic and serum IL-22 expression levels are significantly elevated
[86]. It has been confirmed that through the STAT3 pathway, IL-22 stimulates the proliferation of liver stem/progenitor cells (LPCs), cells participating in liver inflammatory responses
[86]. Moreover, IL-22 production in hepatitis B patients is positively related to the HBV load
[87]. By increasing CXCL9 and CXCL10 expression on hepatic stellate cells (HSCs), IL-22 also promotes inflammatory cell accumulation in the liver, leading to increased liver damage and hepatic fibrosis
[88]. As a result, IL-22 may be both proinflammatory and profibrogenic in hepatitis B. In addition, elevated circulating levels of Th22 correlated with the severity of HBV-associated acute–chronic liver failure (HBV-ACLF), suggesting that Th22 cells are a negative predictor of prognosis in HBV-ACLF
[89]. Nevertheless, according to Kong et al., IL-22 can activate STAT3 and p53 to induce senescence in HSCs
[90]. IL-22 levels also negatively correlate with the development of liver fibrosis
[91]. Therefore, the proinflammatory or protective effect of IL-22 during HBV infection may vary according to different disease states
[92][93][92,93].
For hepatitis C, IL-22 mRNA and Th22 cell levels were elevated in the livers of patients with chronic hepatitis C (CHC) compared to controls
[84][93][94][84,93,94], indicating that Th22 cells may be recruited to the liver by intrahepatic chemokines
[94]. Furthermore, IL-22 induces the proliferation of HSCs, and the Th22 cell level is positively related to the progression of CHC to cirrhosis, suggesting that Th22/IL-22 facilitates HCV-related liver fibrosis
[95][96][95,96]. However, an in vitro study confirmed that two variants of the gene encoding IL-22BP are correlated with HCV-mediated liver fibrosis and cirrhosis. It has also been proven that high production of IL-22 is correlated with protective immune responses to hepatitis C
[51]. Furthermore, during the progression of hepatitis C to cirrhosis, the ratio of IL-22BP/IL-22 increases with the stage of liver fibrosis and peaks at the time of cirrhosis
[97]. Hence, the administration of IL-22BP inhibitors, such as IL-18, prostaglandin E2 (PGE2), NLRP3 and NLRP6 inflammasomes
[98][99][100][98,99,100], may be a promising therapy for liver fibrosis
[51]. This conclusion was questioned by Wu et al.
[101]. They stated that the correlation between IL-22 and protective responses in vitro may not be available in vivo because physiological IL-22 levels in the liver cannot be accurately measured in vitro
[101], which means that the exact role of IL-22 in hepatitis C remains controversial.
2.1.4. Influenza
Influenza is a highly infectious disease that primarily invades the respiratory system. By attacking lung epithelial cells, influenza viruses can destroy the pulmonary epithelial barrier and lead to abnormal gas exchange and pulmonary effusion
[102]. After infection with influenza A virus (IAV), IL-22 levels in the lung tissues of patients are significantly increased and can gradually return to normal as the disease improves
[103][104][105][103,104,105]. Moreover, the lung injury caused by H1N1 IAV is more severe in IL-22
-/- mice than in controls
[57]. These findings suggest that IL-22 may have a protective role during influenza infection. A study showed that some IAV-infected mice can inhibit IL-22 production by producing type I interferons (I-IFN), and they were at a higher risk of secondary infection with
S. aureus [106]. In mice infected with IAV, IL-22 protects the integrity of the epithelial barrier and inhibits secondary infection by inducing antimicrobial peptides and intercellular junction proteins expressed in the respiratory epithelium
[107]. Tight junction proteins promote fluid efflux to alleviate pulmonary effusion in influenza A patients
[108][109][108,109]. In addition, IL-22 has also been proven to inhibit IAV-induced lung epithelial cell necrosis, suppress inflammatory responses and promote bronchial epithelial cell regeneration
[104][105][110][104,105,110]. Since IL-22 can be blocked by IL-22BP, IL-22BP inhibitors have been assumed to be effective at improving influenza prognosis
[54]. Nevertheless, in contrast to the upregulated IL-22 expression in patients with a mild infection, the IL-22 levels were decreased remarkably in patients with severe influenza A
[111]. Accordingly, IL-22 protects the respiratory system in IAV-infected patients and can be dysregulated with disease progression.
2.1.5. Acute Viral Myocarditis
Acute viral myocarditis (AVMC) is nonspecific interstitial myocardial inflammation. Coxsackievirus B3 (CVB3) infection is the major cause
[112]. Compared to the controls, circulating Th22 cells and IL-22 were significantly upregulated in mice infected with CVB3
[113]. Furthermore, in CVB3-infected mice, the anti-IL-22 antibody reduced the antiviral IFN-γ production while increasing the levels of proinflammatory cytokines such as IL-17, IL-6 and TNF-α
[113]. Consequently, the antibody resulted in the deterioration of AVMC, demonstrating that Th22/IL-22 can regulate the expression of cytokines and improve antiviral activity and prognosis during CVB3 infection. If left untreated, AVMC can progress to dilated cardiomyopathy (DCM)
[112]. Using animal models of CVB3-induced chronic myocarditis and DCM, studies have revealed that Th22/IL-22 also has a protective role in chronic viral myocarditis and that IL-22 can inhibit cardiac fibrosis
[114]. Therefore, Th22/IL-22 may be a promising target for treating coxsackievirus-induced acute viral myocarditis, chronic viral myocarditis, and DCM. Nevertheless, in IL-17A
-/- mice with AVMC, IL-22 neutralization contributes to improving acute myocarditis while increasing viral replication at the same time
[115]. This result suggests that when IL-17A is absent, IL-22 can exacerbate the progression of AVMC and inhibit CVB3 replication.
2.1.6. Other Viral Infections
Hand, foot, and mouth disease (HFMD) occurs mainly in children infected with coxsackievirus A16 (CV-A16) or enterovirus 71 (EV-71), leading to characteristic herpes on the hand, foot, mouth, and buttock
[116]. In the acute phase of EV-71-induced HFMD, levels of both circulating Th22 cells and IL-22 are higher than in the convective phase. In addition, HFMD patients complicated with viral encephalitis have higher levels of IL-22 than patients with HFMD alone
[117]. This result suggests that Th22/IL-22 may be crucial in the progression of HFMD
[118]. Strong bronchitis in infants
[119][120][119,120] and pneumonia in elderly or low-resistance patients
[121] are often caused by respiratory syncytial virus (RSV) infection. The robust Th22 response during the acute phase of RSV infection predicts prolonged hospitalization, indicating a negative effect of Th22/IL-22 on RSV infection
[122]. Nevertheless, contrary to the pathogenic effect of endogenous IL-22, exogenous IL-22 activates the STAT3 pathway in RSV-infected cells to promote apoptosis and inhibit viral replication
[58]. Therefore, the administration of IL-22 may be effective in RSV therapy. Warts are a contagious skin disease primarily caused by human papillomavirus (HPV)
[123]. In comparison with healthy controls, patients with warts produced more IL-22 in the serum, and their IL-22 levels were positively related to the wart counts, suggesting that IL-22 is involved in the immune response against HPV
[124]. Yellow fever (YF) is endemic in the tropics and is characterized by acute fever, jaundice, and proteinuria
[125][126][125,126]. It is typically induced by infection with yellow fever virus (YFV)
[125]. Compared with healthy controls, IL-22 production by liver parenchymal cells from YF patients is significantly upregulated
[127]. In addition, Mendes et al. speculated that IL-22 participates in M2 macrophage-mediated organ repair and the immune escape mechanism of YFV
[127].
Consequently, Th22/IL-22 has both a protective and a proinflammatory role in viral infections, which is greatly affected by factors such as the subtype of virus, the severity of infection and the presence of IL-17A.
2.2. Th22 Cells in Bacterial Infections
2.2.1. Mycobacterium tuberculosis
Mycobacterium tuberculosis (MTB) is a common pathogen that is primarily transmitted via the respiratory tract
[128]. Compared to healthy controls, TB patients had lower levels of IL-22 and IL-22
+ T cells in their plasma
[129][130][129,130]. Th22 cells are the main IL-22 producers during
MTB infection, but other subsets, such as Th1 cells, CD8+ T cells, and NKT cells, also secrete IL-22
[131][132][133][131,132,133]. In addition, the bronchoalveolar lavage fluid (BALF) of pulmonary TB patients contains a large amount of IL-22 at significantly higher levels than the corresponding plasma
[134]. IL-22 was also found to be abundant in both TB-induced pleural and pericardial effusions
[135]. Therefore, it illustrated a possible aggregation of IL-22-producing cells in the disease sites of TB patients.
MTBs in pulmonary TB granulomas have been shown to recruit IL-22
+ T cells
[136]. During
MTB infection, since IL-22R is mainly expressed on the surfaces of macrophages in tuberculous granulomas, it may also contribute to Th22 cell accumulation
[132][137][132,137]. Furthermore, in tuberculous pleurisy, the accumulation of Th22 cells at the disease site was associated with the chemotactic effect of cytokines in tuberculous pleural effusion (TPE) and the pleural mesothelial cell (PMC)-expressed chemokines CCL20, CCL22 and CCL27
[138]. PMCs also promote Th22 proliferation and differentiation by presenting
MTB antigens
[138].
MTB infection in humans can result in asymptomatic specific immune responses, latent tuberculosis (TB) or active TB
[128]. Bunjun et al. observed a high level of IL-22 in latent
MTB patients stimulated by
MTB antigens, and IL-22 accounted for the largest proportion of responsive CD4+ responses
[131]. Compared to wild-type controls, IL-22
-/- mice showed higher susceptibility to
MTB HN878 and a higher bacterial load in the lungs during the chronic phase of
MTB infection
[132]. In addition, the pulmonary
MTB load at the early stage is significantly increased in IL-22-deficient mice
[139]. Therefore, IL-22 is required in both adaptive and innate immune responses against
MTB.
Research has shown that in response to stimulation by
MTB, Th22 cells can evolve into membrane-bound IL-22
+ (mIL-22
+) Th22 cells to extend the half-life of IL-22
[137]. More importantly, mIL-22 binds to IL-22R on infected macrophages to inhibit intracellular
MTB replication
[137]. IL-22 also enhances the expression of CCL2 on epithelial cells to stimulate the pulmonary recruitment of macrophages
[132]. In
MTB-infected phagocytes, IL-22 modulates the expression of Rab7 and Rab14 by upregulating the production of calgranulin A
[140][141][140,141]. Rab7 and Rab14 subsequently inhibit intracellular
MTB replication and promote phagosome maturation and fusion
[140][141][140,141]. Furthermore, IL-22 was found to stimulate TNF-α production by IL-22R
+ macrophages
[132]. TNF-α can promote macrophage activation and
MTB control directly. Additionally, the level of IL-22 in tuberculous pericardial effusion is positively correlated with MMP-9, an enzyme capable of degrading the extracellular matrix
[135]. Since the peripheral level of MMP-9 is associated with the severity of tuberculosis
[142], IL-22 might also regulate MMP-9 expression to control
MTB infection
[135]. Moreover, IL-22 can promote the proliferation and recovery of pleural mesothelial cells (PMCs) and the closure of PMC layers
[138], which facilitates protection of PMCs from tuberculosis-induced damage. Antimicrobial proteins induced by IL-22 in the chronic phase, such as RegIIIγ, Lcn2 and calgranulin A, ensure the structural and functional integrity of the pulmonary epithelial barrier
[132]. IL-22 is also involved in the formation of protective lymphoid follicular structures, called inducible bronchus-associated lymphoid tissues (iBALT), through the CXCR5–CXCL13 axis
[139]. In patients infected with
multidrug-resistant MTB (MDR-TB), decreased Th22 cell responses are associated with high sputum bacterial loads and severe lung lesions, suggesting that Th22 cells influence the antimicrobial capacity of TB patients
[129]. Moreover, fewer T cells that release PD-1 or CD57 were found in multidrug-resistant TB patients with high levels of Th22 cells
[129]. CD57 and programmed cell death 1 (PD-1) are markers of cellular senescence, so this result suggests that the Th22-induced anti-tuberculosis immune response may also be related to decreased T-cell senescence. Therefore, Th22 cells and IL-22 inhibit the progression of TB by protecting the pulmonary epithelial barrier and regulating
MTB-specific immune responses.
2.2.2. Citrobacter Rodentium
Since
Citrobacter rodentium-infected mice closely resemble human infectious colitis, they are commonly used in colitis-associated scientific research
[143].
C. rodentium infection in mice dramatically induces the expression of IL-22 in the intestinal mucosa
[143]. Studies have confirmed that during
C. rodentium infection, early IL-22 production is primarily dependent on ILC3s, whereas IL-22 derived from Th22 cells predominates at a later stage
[144]. Th17 cells are also involved in IL-22 production
[144]. Further experiments injected purified Th22 cells, Th17 cells or ILC3s into
C. rodentium-infected mice deficient in IL-22. Compared to the other two subsets, mice injected with Th22 cells had a higher survival rate, reflecting that Th22/IL-22 is critical for the host’s immune response to
C. rodentium [144]. Moreover, when the STAT3 gene was mutated,
C. rodentium-infected mice showed a loss of Th22-induced immune defense, including impairment of the intestinal epithelial barrier, reduction of antimicrobial peptides, and increased dissemination of pathogenic bacteria
[143]. This confirms that STAT3 is indispensable for the protective role of IL-22 during
C. rodentium infection. In addition to STAT3, Th22/IL-22-related effects are dependent on T-bet and AhR
[143][144][143,144]. Furthermore, IL-22 can induce colonic epithelial cells to express the antibacterial proteins RegIIIβ and RegIIIγ, both of which play a direct bactericidal role in
C. rodentium infection
[37]. Researchers have shown that antibiotic treatment reduces RegIIIγ production in mice, but oral administration of lipopolysaccharide (LPS) can restore it
[145]. This indicates that LPS, the principal ingredient of the Gram-negative bacterial outer membrane, is key to the expression of RegIIIγ
[145]. The addition of the Toll-like receptor (TLR)-5 agonist flagellin or the binding between TLR and its ligand MyD88 has been revealed to initiate the production of IL-23 by DCs and promote IL-23 to upregulate IL-22 production and RegIIIγ expression
[146][147][146,147]. As a consequence, the host defense against
C. rodentium induced by Th22 cells is dependent on STAT3 activation and antibacterial protein expression.
2.2.3. Streptococcus pneumoniae
Streptococcus pneumoniae is a Gram-positive bacterium that colonizes the nasopharynx and initiates respiratory inflammation
[148]. According to previous studies, IL-22 expression was rapidly upregulated in the lungs of mice suffering from pneumococcal pneumonia
[149]. Moreover, the lung bacterial load was significantly higher in mice with hepatic IL-22R1 deficiency than in controls
[149]. Hence, the IL-22-induced downstream cascade is critical for inhibiting
S. pneumoniae replication. IL-22 was confirmed to promote the clearance of
S. pneumoniae by upregulating complement factor C3’s expression in the liver and strengthening the binding of C3 and
S. pneumoniae in the serum
[149]. In addition, mice deficient in IL-22RA2 (the gene encoding IL-22BP) showed reduced susceptibility to
S. pneumoniae and prolonged survival after infection
[55]. Further analysis revealed that in the lungs of IL-22RA2
-/- mice, oxidative phosphorylation-related genes were reduced in expression. Downregulated oxidative phosphorylation led to an increase in glycolysis and promoted a shift in macrophage phenotypes toward a proinflammatory phenotype, which ultimately upregulated host resistance to pneumococcal pneumonia. Consequently, IL-22 and IL-22BP coregulate antimicrobial immunity against
S. pneumoniae, suggesting that IL-22RA2 may be an effective target for the treatment of pneumococcal pneumonia
[55].
2.2.4. Helicobacter pylori
Helicobacter pylori is Gram-negative and colonizes the gastric mucosa, causing persistent gastric inflammation, peptic ulcers or even gastric cancer
[150][151][150,151]. Zhuang et al. demonstrated that Th22 cells are proinflammatory in
H. pylori-triggered gastritis
[152]. Upon
H. pylori infection, IL-23-induced Th22 cells are rapidly enriched in the gastric mucosa and secrete IL-22, which enhances CXCL2 production by gastric epithelial cells
[152]. CXCL2 subsequently bound to its corresponding receptors and resulted in the migration of myeloid-derived suppressor cells (MDSCs) toward the gastric epithelium. In response to IL-22 induction, MDSCs produce the proinflammatory factors calgranulin A (S100A8) and S100A9 and directly inhibit the development of Th1 cells, leading to gastritis progression
[152]. Moreover, through activation of the ERK pathway, IL-22 and Helicobacter pylori synergistically promote the production of matrix metalloproteinases (MMPs), particularly MMP-10, in gastric epithelial cells
[153]. MMP-10 can aggravate bacterial colonization by inhibiting the production of antimicrobial peptides in the gastric mucosa. In addition, MMP-10 also induces gastric epithelial cells to secrete the chemokine CXCL16, which recruits CD8+ T cells to the gastric mucosa and exacerbates the inflammatory response. However, when IL-22 acts synergistically with IL-17A, it stimulates antimicrobial peptide expression that protects the body from
H. pylori infection
[154]. Moreover, it has been proposed that due to the compensatory effect of other cytokines, IL-22 deficiency does not affect the susceptibility of mice to
H. pylori or their mortality after infection
[154], which contradicts the previous conclusion. According to a paper published in 2019, the discrepancy may be attributed to their choice of different mouse species and
H. pylori strains, which closely influence the observed immune response
[150]. Therefore, the role of IL-22/Th22 in
H. pylori infection remains to be further discussed.
2.2.5. Other Bacterial Infections
Clostridium difficile typically causes pseudomembranous colitis or bacterial diarrhea in immunocompromised individuals by facilitating the translocation of enteropathogenic bacteria
[155][156][155,156]. Compared to the wild-type group, IL-22-deficient mice showed increased mortality after infection with
C. difficile [156]. During
C. difficile infection, induced IL-22 upregulates the expression of acute-phase proteins and C3 in the liver and intestine
[156]. C3 can be deposited on the surface of enteropathogenic bacteria and enhance the bactericidal activity of neutrophils
[156]. Moreover, IL-22-induced glycosylation of host N-linked glycans can promote the growth of
Phascolarctobacterium spp. [157].
Phascolarctobacterium spp. is a succinate-consuming commensal bacterium that competes with
C. difficile for energy, preventing the growth and colonization of
C. difficile in the intestine. Infection with
Klebsiella pneumoniae is often associated with healthcare-related pneumonia or sepsis
[158]. Compared with uninfected mice, IL-22 levels in the lungs of mice infected with
K. pneumoniae were upregulated
[44]. The upregulation of IL-22 can reverse the decreased epithelial barrier stability and deteriorated pulmonary inflammation caused by IFN-λ
[159]. IL-22 also induces the expression of defense genes such as lipocalin 2 to suppress
K. pneumoniae [160]. Lipocalin 2 can sequester iron from bacteria to limit their growth
[160][161][160,161]. Moreover, IL-22 stimulates the lung tissues of
K. pneumoniae-infected mice to express CCL17 and CCL20
[44], which are ligands of CCR4 and CCR6
[17], leading to the accumulation of Th22 cells in the lungs.
Pseudomonas aeruginosa is an opportunistic bacterium that can cause an overwhelming local immune response and consistently lead to acute respiratory distress syndrome (ARDS)
[162]. The level of IL-22 was transiently increased in
P. aeruginosa-infected mice, and additional administration of IL-22 reduced their pulmonary damage by downregulating local neutrophil infiltration
[163]. IL-22 also induces pulmonary IFN-λ production, preventing the release of inflammatory mediators such as IL-1β
[164]. Notably, both protease IV secreted by
P. aeruginosa [165] and serine protease-3 secreted by neutrophils
[166] can degrade IL-22. Therefore, they facilitate the immune escape of
P. aeruginosa and can result in pulmonary bacterial colonization and the continued deterioration of respiratory function. Consequently, Th22/IL-22 is protective during
C. difficile,
K. pneumoniae and
P. aeruginosa infections. However,
Salmonella enterica serotype Typhimurium colonizes the gastrointestinal mucosa and typically causes inflammatory diarrhea
[167]. The iroBCDE iroN gene cluster
[168] and zinc transporter (ZnuABC)
[169] in
S. Typhimurium enable them to escape from IL-22-mediated metal chelation, called nutritional immunity
[170]. Since IL-22 is primarily secreted by ILC3s rather than T cells during
S. Typhimurium infection
[171], more detailed mechanisms will not be discussed
in th
ereis review.
Therefore, Th22/IL-22 exerts a crucial effect on regulating the immune response to viral and bacterial infections. Based on the above, cytokines such as TNF-α, IFN-λ, IL-10 and IL-23; transcription factors such as T-bet and AhR; and chemokines such as CCL2, CCL17 and CXCL13 have been shown to have a protective role in the Th22/IL-22 axis. Other effector molecules, including C3, CRP, MMP-9, Rab7, Rab14, the TLR-5 agonist flagellin and the antibacterial proteins RegIIIβ and RegIIIγ, are also involved in IL-22-induced anti-infectious immunity. Moreover, the matrix metalloproteinase MMP-10; the antimicrobial protein S100A9; and chemokines such as CXCL9, CXCL10, CXCL2 and CXCL16 are risk factors for the Th22/IL-22 downstream signaling system. Interestingly, STAT3 and the antimicrobial protein calgranulin A can be both protective and deleterious in the Th22/IL-22 axis, depending on the type of infectious disease. Notably, IL-22 in infectious illnesses is also derived from ILC3s, Th1 cells, Th17 cells and NKT cells, indicating that the interregulation between different cells ultimately contributes to the development of diseases. This suggests the need for more research on the underlying mechanisms of Th22/IL-22 in infectious diseases.