Salt (NaCl) intake is processed by receptors in the tongue and digestive system, which transmit the information to the nucleus of the solitary tract via a neural pathway (chorda tympani/vagus nerves) and to circumventricular organs, including the subfornical organ and area postrema, via a humoral pathway (blood/cerebrospinal fluid). Circuits are formed that stimulate or inhibit homeostatic sodium (Na) intake involving participation of the parabrachial nucleus, pre-locus coeruleus, medial tuberomammillary nuclei, median eminence, paraventricular and supraoptic nuclei, and other structures with reward properties such as the bed nucleus of the stria terminalis, central amygdala, and ventral tegmental area. Finally, the kidney uses neural signals (e.g., renal sympathetic nerves) and vascular (e.g., renal perfusion pressure) and humoral (e.g., renin–angiotensin–aldosterone system, cardiac natriuretic peptides, antidiuretic hormone, and oxytocin) factors to promote Na excretion or retention and thereby maintain extracellular fluid volume.
- sodium homeostasis
- hypernatremia
- hyponatremia
1. Introduction
2. Behavioral Mechanism: Salt Intake
The Na ion [Na+] is an essential micronutrient that cannot be generated by endogenous processes and must be obtained through the diet [9]. The need to maintain an appropriate Na balance has led living organisms to develop mechanisms that identify the presence of Na the instant it enters the body [10]. The first level of detection is located in the oral cavity, from where it accesses the digestive system and is absorbed in the blood.2.1. Detection and Processing in the Oral Cavity
Na is detected by the taste system within the oral cavity. The mineral generates a salty taste that guides and maintains its consumption [11][12][13][14]. The gustatory stimulus used as prototype to study the perception of this taste was table salt (NaCl), physiologically the most important salt in the diet, which induces the purest salty taste [11][15], and biologically the most relevant to maintain biochemical balance [16]. In mammals, gustatory receptors are observed throughout the oral cavity, especially on the tongue, where they are located on protuberances designated papillae. There are four types of papillae according to their shape and localization: fungiform (anterior two-thirds of the tongue), foliate (side toward the back), circumvallate (on the back), and filiform (anterior two-thirds of the tongue). Cells that detect NaCl are mainly located in fungiform papillae [12][17]. In general, gustatory papillae can contain several hundreds of taste buds, onion-shaped organs (Figure 1) that house taste receptor cells (50–100 cells each), i.e., the cells responsible for detecting and transducing taste stimuli (transforming chemical signals into neural activity). In the taste bud, these cells are mechanically held together by tight junctions that separate apical areas from the remaining basolateral region, forming an almost impermeable barrier. Only the apical membrane above the tight junctions (2–3% of the total membrane area of the cell) is exposed to saliva, while the basolateral area remains in a relatively constant interstitial milieu [12][13][14][18].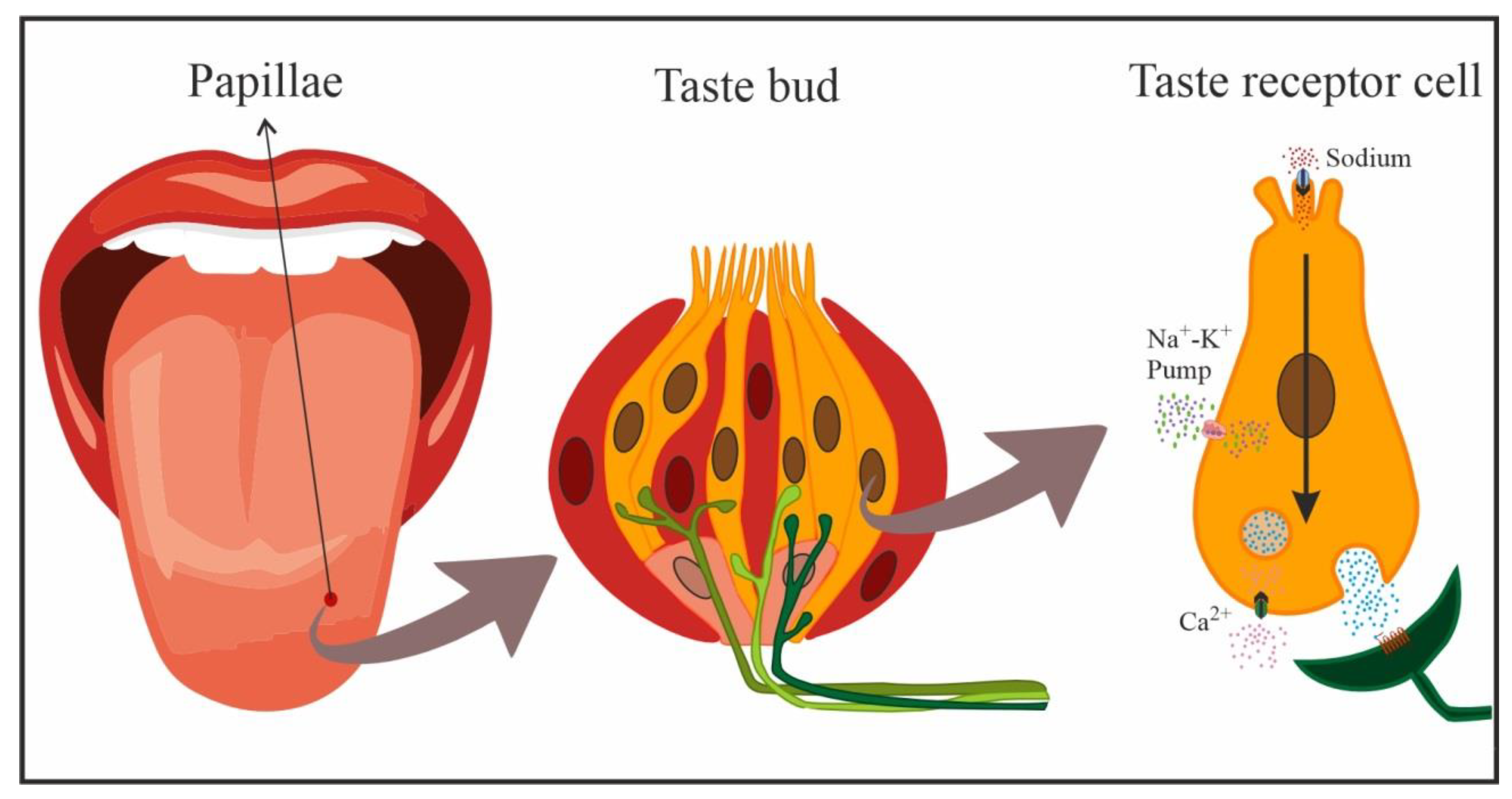
2.2. Sodium Detection and Processing in the Gastrointestinal System
Various studies have demonstrated that the most caudal levels of the digestive system are also prepared for Na detection (visceral sensory signals) [29][30][31][32][33]. Information on the entry of Na can be transmitted from the gastrointestinal system to the central nervous system (CNS) in a rapid manner, via neural mechanisms, or more slowly, via the circulatory system (humoral pathway) [29][32][33][34][35][36]. The neural pathway is vagal in nature [37], and Na+ detection by subdiaphragmatic vagal afferents is well documented [33][38]. The presence of NaCl-sensitive receptors has been demonstrated in the gastrointestinal mucosa [39][40][41][42] and at hepatic level [43][44]. These receptors appear to be connected to small-diameter fibers, mostly unmyelinated vagal fibers that respond to the presence of hypertonic NaCl with a mean latency of around 10–20 s [39][40][41][42]. The first relay of visceral-vagal information from post-oral segments of the digestive system takes place in caudal segments of the nucleus of the solitary tract (NST), from which it is remitted to other brain structures [27][45][46]. Na absorption into the circulatory system largely takes place in the distal small bowel and the colon [47][48][49]. In these digestive segments, Na+ is taken up through absorptive cells of the epithelium, which connect on one side with the intestinal lumen (apical region, brush border) and on the other (basolateral region) with blood and lymphatic capillaries. In the small intestine, these cells are organized into digitiform projections or intestinal villi that significantly augment (around 600-fold) the absorption surface area; however, this organization is less marked towards distal ends of the intestine and is virtually absent at the most distal region of the colon [50]. Na absorption is transcellular in the intestine. In a first step, the ion penetrates from the intestinal lumen into the cytoplasm of the absorptive epithelial cell following electrochemical gradients and strongly assisted by Na-dependent transporters (e.g., Na+/glucose and Na+/cotransporter amino acids or Na–hydrogen exchangers (NHEs), or by Na reuptake through ENaCs mainly present in the distal colon [50][51]. Once within the epithelial cells, Na+ is actively transported towards paracellular spaces by the Na+/K+ ATPase pump in the basal and lateral membranes of the cells, avoiding retrograde diffusion, from where it enters the bloodstream [47][48][50]. The modality of Na+ absorption and the intestinal segment in which it occurs mainly depends on the circumstances. In this way, it appears to occur preferentially in the small intestine after meals, coupled with nutrient absorption, nutrient-dependent Na absorption, Na+/glucose, and Na+/cotransporter amino acids. In contrast, it largely takes place in the ileum and colon through NHE or ENaCs during interdigestive periods [50][52]. Under Na deficiency conditions, hormones such as aldosterone can increase Na uptake in the intestinal epithelium, stimulating the expression of the transporters that participate in its absorption (especially at colon level). Consequently, there is virtually no Na loss under these circumstances, with <0.5% of the intestinal content of Na being excreted via the feces under normal conditions [47][51][53][54]. Once in the circulatory system, Na is widely distributed throughout the organism, accessing circumventricular brain structures, which lack a blood–brain barrier, via fenestrated capillaries [55][56], and accessing cerebrospinal fluid (CSF) via ion transporters (e.g., ENaC) in epithelial cells of the choroid plexus [57][58][59][60]. Blood and CSF both have a [Na+] concentration of 144 mOsm/L [61].References
- Fessler, D.M.T. An Evolutionary Explanation of the Plasticity of Salt Preferences: Prophylaxis against Sudden Dehydration. Med. Hypotheses 2003, 61, 412–415.
- Leshem, M. Salt Need Needs Investigation. Br. J. Nutr. 2020, 123, 1312–1320.
- Schulkin, J. Sodium Hunger: The Search for a Salty Taste; Cambridge University Press: Cambridge, UK, 1991; ISBN 978-0-521-35368-7.
- Bie, P. Mechanisms of Sodium Balance: Total Body Sodium, Surrogate Variables, and Renal Sodium Excretion. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 315, R945–R962.
- Strazzullo, P.; Leclercq, C. Sodium. Adv. Nutr. 2014, 5, 188–190.
- Verbalis, J.G.; Stricker, E.M. Neuroendocrine Regulation of Fluid Intake and Homeostasis. In Neuroendocrinology in Physiology and Medicine; Conn, P.M., Freeman, M.E., Eds.; Humana Press: Totowa, NJ, USA, 2000; pp. 317–334. ISBN 978-1-61737-153-0.
- Mahía, J.; Bernal, A. Animal Models for Diabetes Insipidus. Handb. Clin. Neurol. 2021, 181, 275–288.
- Noda, M.; Matsuda, T. Central Regulation of Body Fluid Homeostasis. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2022, 98, 283–324.
- Lundy, R.F. Potential Mechanisms for Functional Changes in Taste Receptor Cells Following Sodium Deficiency in Mammals. Neurosci. Biobehav. Rev. 1998, 23, 103–109.
- McCaughey, S.A.; Scott, T.R. The Taste of Sodium. Neurosci. Biobehav. Rev. 1998, 22, 663–676.
- Bigiani, A. Does ENaC Work as Sodium Taste Receptor in Humans? Nutrients 2020, 12, 1195.
- Diepeveen, J.; Moerdijk-Poortvliet, T.C.W.; van der Leij, F.R. Molecular Insights into Human Taste Perception and Umami Tastants: A Review. J. Food Sci. 2022, 87, 1449–1465.
- Lindemann, B. Taste Reception. Physiol. Rev. 1996, 76, 719–766.
- Wu, A.; Dvoryanchikov, G.; Pereira, E.; Chaudhari, N.; Roper, S.D. Breadth of Tuning in Taste Afferent Neurons Varies with Stimulus Strength. Nat. Commun. 2015, 6, 8171.
- DeSimone, J.A.; Lyall, V. Taste Receptors in the Gastrointestinal Tract III. Salty and Sour Taste: Sensing of Sodium and Protons by the Tongue. Am. J. Physiol. Gastrointest. Liver Physiol. 2006, 291, G1005–G1010.
- Miyamoto, T.; Fujiyama, R.; Okada, Y.; Sato, T. Acid and Salt Responses in Mouse Taste Cells. Prog. Neurobiol. 2000, 62, 135–157.
- Oakley, B.; Witt, M. Building Sensory Receptors on the Tongue. J. Neurocytol. 2004, 33, 631–646.
- Travers, J.B.; Travers, S.P.; Norgren, R. Gustatory Neural Processing in the Hindbrain. Annu. Rev. Neurosci. 1987, 10, 595–632.
- Kinnamon, S.C.; Finger, T.E. Recent Advances in Taste Transduction and Signaling. F1000Res 2019, 8, F1000 Faculty Rev-2117.
- Chandrashekar, J.; Kuhn, C.; Oka, Y.; Yarmolinsky, D.A.; Hummler, E.; Ryba, N.J.P.; Zuker, C.S. The Cells and Peripheral Representation of Sodium Taste in Mice. Nature 2010, 464, 297–301.
- Herness, M.S. Aldosterone Increases the Amiloride-Sensitivity of the Rat Gustatory Neural Response to NaCl. Comp. Biochem. Physiol. Comp. Physiol. 1992, 103, 269–273.
- Shigemura, N.; Iwata, S.; Yasumatsu, K.; Ohkuri, T.; Horio, N.; Sanematsu, K.; Yoshida, R.; Margolskee, R.F.; Ninomiya, Y. Angiotensin II Modulates Salty and Sweet Taste Sensitivities. J. Neurosci. 2013, 33, 6267–6277.
- Frank, M.E.; Contreras, R.J.; Hettinger, T.P. Nerve Fibers Sensitive to Ionic Taste Stimuli in Chorda Tympani of the Rat. J. Neurophysiol. 1983, 50, 941–960.
- McCaughey, S.A. Characterization of Mouse Chorda Tympani Responses Evoked by Stimulation of Anterior or Posterior Fungiform Taste Papillae. Neurosci. Res. 2019, 141, 43–51.
- Spector, A.C.; Grill, H.J. Salt Taste Discrimination after Bilateral Section of the Chorda Tympani or Glossopharyngeal Nerves. Am. J. Physiol. 1992, 263, R169–R176.
- Stricker, E.M.; Gannon, K.S.; Smith, J.C. Thirst and Salt Appetite Induced by Hypovolemia in Rats: Analysis of Drinking Behavior. Physiol. Behav. 1992, 51, 27–37.
- Altschuler, S.M.; Bao, X.; Bieger, D.; Hopkins, D.A.; Miselis, R.R. Viscerotopic Representation of the Upper Alimentary Tract in the Rat: Sensory Ganglia and Nuclei of the Solitary and Spinal Trigeminal Tracts. J. Comp. Neurol. 1989, 283, 248–268.
- Hamilton, R.B.; Norgren, R. Central Projections of Gustatory Nerves in the Rat. J. Comp. Neurol. 1984, 222, 560–577.
- Arnedo, M.; Gallo, M.; Agüero, A.; Puerto, A. Effects of Medullary Afferent Vagal Axotomy and Area Postrema Lesions on Short-Term and Long-Term NaCl-Induced Taste Aversion Learning. Physiol. Behav. 1990, 47, 1067–1074.
- Burman, A.; Kaji, I. Luminal Chemosensory Cells in the Small Intestine. Nutrients 2021, 13, 3712.
- Sbarbati, A.; Osculati, F. The Taste Cell-Related Diffuse Chemosensory System. Prog. Neurobiol. 2005, 75, 295–307.
- Zafra, M.A.; Prados, M.; Molina, F.; Puerto, A. Capsaicin-Sensitive Afferent Vagal Fibers Are Involved in Concurrent Taste Aversion Learning. Neurobiol. Learn. Mem. 2006, 86, 349–352.
- Zimmerman, C.A.; Huey, E.L.; Ahn, J.S.; Beutler, L.R.; Tan, C.L.; Kosar, S.; Bai, L.; Chen, Y.; Corpuz, T.V.; Madisen, L.; et al. A Gut-to-Brain Signal of Fluid Osmolarity Controls Thirst Satiation. Nature 2019, 568, 98–102.
- Johnson, A.K.; Thunhorst, R.L. The Neuroendocrinology of Thirst and Salt Appetite: Visceral Sensory Signals and Mechanisms of Central Integration. Front. Neuroendocr. 1997, 18, 292–353.
- Mediavilla, C.; Bernal, A.; Mahía, J.; Puerto, A. Nucleus of the Solitary Tract and Flavor Aversion Learning: Relevance in Concurrent but Not Sequential Behavioral Test. Behav. Brain Res. 2011, 223, 287–292.
- Mediavilla, C.; Molina, F.; Puerto, A. Concurrent Conditioned Taste Aversion: A Learning Mechanism Based on Rapid Neural versus Flexible Humoral Processing of Visceral Noxious Substances. Neurosci. Biobehav. Rev. 2005, 29, 1107–1118.
- Contreras, R.J.; Kosten, T. Changes in Salt Intake after Abdominal Vagotomy: Evidence for Hepatic Sodium Receptors. Physiol. Behav. 1981, 26, 575–582.
- Chernigovsky, V.N. The Significance of Interoceptive Signals in the Food Behavior in Animals. In The Internal Environment and Alimentary Behavior; Brazier, M.A.B., Ed.; Brain and Behavior; American Institute of Biological Sciences, University of California: Los Angeles, CA, USA, 1963; pp. 319–349.
- Blackshaw, L.A.; Grundy, D. Effects of 5-Hydroxytryptamine on Discharge of Vagal Mucosal Afferent Fibres from the Upper Gastrointestinal Tract of the Ferret. J. Auton. Nerv. Syst. 1993, 45, 41–50.
- Mei, N. Intestinal Chemosensitivity. Physiol. Rev. 1985, 65, 211–237.
- Mei, N.; Garnier, L. Osmosensitive Vagal Receptors in the Small Intestine of the Cat. J. Auton. Nerv. Syst. 1986, 16, 159–170.
- Zhu, J.X.; Wu, X.Y.; Owyang, C.; Li, Y. Intestinal Serotonin Acts as a Paracrine Substance to Mediate Vagal Signal Transmission Evoked by Luminal Factors in the Rat. J. Physiol. 2001, 530, 431–442.
- Kahrilas, P.J.; Rogers, R.C. Rat Brainstem Neurons Responsive to Changes in Portal Blood Sodium Concentration. Am. J. Physiol. 1984, 247, R792–R799.
- Morita, H.; Yamashita, Y.; Nishida, Y.; Tokuda, M.; Hatase, O.; Hosomi, H. Fos Induction in Rat Brain Neurons after Stimulation of the Hepatoportal Na-Sensitive Mechanism. Am. J. Physiol. 1997, 272, R913–R923.
- Barraco, R.; El-Ridi, M.; Ergene, E.; Parizon, M.; Bradley, D. An Atlas of the Rat Subpostremal Nucleus Tractus Solitarius. Brain Res. Bull. 1992, 29, 703–765.
- Herbert, H.; Moga, M.M.; Saper, C.B. Connections of the parabrachial nucleus with the nucleus of the solitary tract and the medullary reticular formation in the rat. J. Comp. Neurol. 1990, 293, 540–580.
- Hall, J.E.; Guyton, A.C.; Hall, M.E. Tratado de Fisiología Médica; 14th ed.; Elsevier: Barcelona, Spain, 2021; ISBN 978-84-13-82013-2.
- Linz, B.; Saljic, A.; Hohl, M.; Gawałko, M.; Jespersen, T.; Sanders, P.; Böhm, M.; Linz, D. Inhibition of Sodium-Proton-Exchanger Subtype 3-Mediated Sodium Absorption in the Gut: A New Antihypertensive Concept. IJC Heart Vasc. 2020, 29, 100591.
- Spiller, R.C. Intestinal Absorptive Function. Gut 1994, 35, S5–S9.
- Binder, H.J. Movimiento de Fluidos y Electrolitos Intestinales—Boron y Boulpaep.Manual de Fisiología Médica. In Boron & Boulpaep Concise Medical Physiology; Boron, W.F., Boulpaep, E.L., Eds.; Elsevier: Philadelphia, PA, USA, 2021; pp. 476–484. ISBN 978-0-323-65530-9.
- Negussie, A.B.; Dell, A.C.; Davis, B.A.; Geibel, J.P. Colonic Fluid and Electrolyte Transport 2022: An Update. Cells 2022, 11, 1712.
- Afsar, B.; Vaziri, N.D.; Aslan, G.; Tarim, K.; Kanbay, M. Gut Hormones and Gut Microbiota: Implications for Kidney Function and Hypertension. J. Am. Soc. Hypertens. 2016, 10, 954–961.
- Coric, T.; Hernandez, N.; de la Rosa, D.A.; Shao, D.; Wang, T.; Canessa, C.M. Expression of ENaC and Serum- and Glucocorticoid-Induced Kinase 1 in the Rat Intestinal Epithelium. Am. J. Physiol. Liver Physiol. 2004, 286, G663–G670.
- Musch, M.W.; Lucioni, A.; Chang, E.B. Aldosterone Regulation of Intestinal Na Absorption Involves SGK-Mediated Changes in NHE3 and Na+ Pump Activity. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 295, G909–G919.
- Ch’ng, S.S.; Lawrence, A.J. The Subfornical Organ in Sodium Appetite: Recent Insights. Neuropharmacology 2019, 154, 107–113.
- Sladek, C.D.; Armstrong, W.E. Osmotic Control of Vasopressin Release. Trends Neurosci. 1985, 8, 166–169.
- Hiyama, T.Y.; Noda, M. Sodium Sensing in the Subfornical Organ and Body-Fluid Homeostasis. Neurosci. Res. 2016, 113, 1–11.
- Miller, R.L.; Wang, M.H.; Gray, P.A.; Salkoff, L.B.; Loewy, A.D. ENaC-Expressing Neurons in the Sensory Circumventricular Organs Become c-Fos Activated Following Systemic Sodium Changes. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 305, R1141–R1152.
- Solár, P.; Zamani, A.; Kubíčková, L.; Dubový, P.; Joukal, M. Choroid Plexus and the Blood-Cerebrospinal Fluid Barrier in Disease. Fluids Barriers CNS 2020, 17, 35.
- Wang, S.; Liu, J.; Cai, H.; Liu, K.; He, Y.; Liu, S.; Guo, Y.; Guo, L. High Salt Diet Elevates the Mean Arterial Pressure of SLC14α1 Gene Depletion Mice. Life Sci. 2020, 254, 117751.
- Peruzzo, M.; Milani, G.P.; Garzoni, L.; Longoni, L.; Simonetti, G.D.; Bettinelli, A.; Fossali, E.F.; Bianchetti, M.G. Body Fluids and Salt Metabolism—Part II. Ital. J. Pediatr. 2010, 36, 78.