Researchers have studied whether growth factors, cytokines, hormones, neurotransmitters, and local hormones (autacoids) promote the proliferation of hepatic parenchymal cells (i.e., hepatocytes) using in vitro primary cultured hepatocytes. The indicators used for this purpose include changes in DNA synthesis activity, nuclear number, cell number, cell cycle, and gene expression. In addition, the intracellular signaling pathways from the plasma membrane receptors to the nucleus have been examined in detail for representative growth-promoting factors that have been found to promote DNA synthesis and cell proliferation of hepatocytes.
- cytokine
- growth factor
- hepatocyte proliferation
- direct mitogen
- indirect mitogen
- co-mitogen
- autocrine mechanism
- signaling pathway
1. Introduction
2. Classification and Characteristics of Hepatocyte Growth Regulators
Mitogens can be defined as factors that promote the proliferation of hepatocytes by themselves, such as EGF, TGF-α, HGF, platelet-derived growth factor (PDGF), and IGF-I. Co-mitogens, by themselves, do not promote hepatocyte proliferation, but, when used in combination with mitogens, the co-mitogens enhance the activity of the mitogens, and these include adrenergic α- and β-agonists and glucagon. In addition, there are many indirect mitogens, such as TNF-α, IL-1β, prostaglandin (PG) E2, 5-HT, and GH, which indirectly promote hepatocyte proliferation by secreting autocrine factors. In contrast, some factors, such as transforming growth factor-β1 (TGF-β1) and glucocorticoids, strongly suppress mitogen-induced hepatocyte proliferation (i.e., they are inhibitory factors).
2.1 Mitogens
2.1.1 Direct Mitogens
EGF
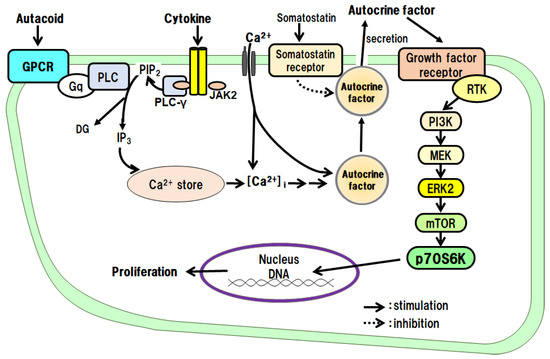
EGF is a protein with a molecular weight of approximately 6 kDa, and it has three disulfide bonds. EGF binds to the EGF receptor (EGFR) and activates intracellular signaling pathways by phosphorylating the receptor tyrosine kinase (RTK); the EGF receptor is phosphorylated in vivo 30–60 min after PHx [3]. The proliferative effects of EGF on hepatocytes have been examined in culture. The results have shown that EGF alone promotes DNA synthesis and the proliferation of hepatocytes. It has been suggested that RTK, extracellular signal-regulated kinase (ERK), and the mammalian target of rapamycin (mTOR) are involved in the signaling [8][13]. Noradrenaline and an adrenergic β2 receptor agonist, on their own, do not promote hepatocyte proliferation, but these agents enhance the growth-promoting effects of EGF. Thus, there is crosstalk between the adrenergic β2 receptor-mediated signaling system and the EGF receptor-mediated signaling system (Figure 1).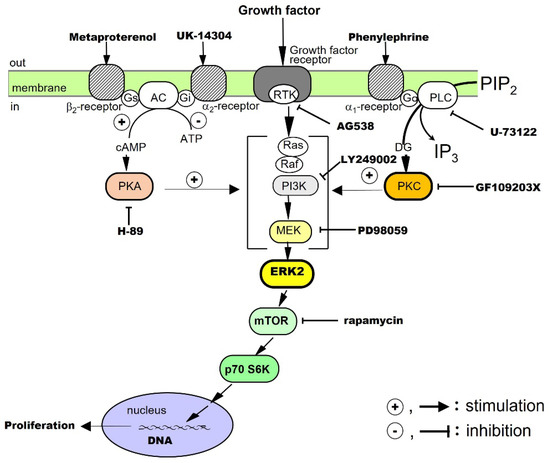
TGF-α
TGF-α is a protein with a molecular weight of approximately 5.5 kDa, which has high amino acid homology with EGF and binds to EGFR/ErbB1. After receptor dimerization, TGF-α activates RTK, which, in turn, activates the mitogen-activated protein (MAP) kinase cascade via Smad, an adaptor protein, for intracellular signal transduction [9][35]. TGF-α, a cytokine produced by keratinocytes and various cancer cells, directly promotes DNA synthesis and the proliferation of hepatocytes alone in culture [10][19]. It is also an autocrine factor secreted by hepatocytes in response to several cytokines (see below). The effects of TGF-α on hepatocytes have been investigated in culture. The results have shown that TGF-α alone promotes DNA synthesis and the proliferation of hepatocytes and that RTK, phosphatidylinositol-3 kinase (PI3K), ERK, and mTOR are involved in its signal transduction. In addition, adrenergic α1 receptor agonists enhance the effects of TGF-α, suggesting crosstalk with adrenergic α1 receptor-mediated signaling pathways (Figure 1).HGF
HGF is a protein consisting of 728 amino acids and a heterodimeric structure, with a heavy chain of approximately 60 kDa and a light chain of 3.5 kDa. It is a cytokine produced by stellate cells and endothelial cells in the liver. Its receptor is cMet, which dimerizes and activates the RTK in the intracellular domain to mediate the action of the HGF [11][12][13][36,37,38]. The proliferative effect of HGF on hepatocytes has been examined in culture. The results have shown that HGF alone promotes DNA synthesis and the proliferation of hepatocytes and that RTK, PI3K, ERK, and mTOR are involved in its signaling [14][20]. ResearchWers have also found that both adrenergic α1 and β2 receptor agonists potentiate the growth-promoting effects of HGF by crosstalk (Figure 1).PDGF
PDGF belongs to the PDGF/VEGF family. Its molecular weight is approximately 30 kDa, and its A and B chains form homo- and hetero-dimeric structures (PDGF-AA, PDGF-BB, and PDGF-AB), which activate RTK for intracellular signal transduction [15][39]. It was first isolated from platelets, but it is also produced by macrophages, vascular endothelial cells, smooth muscle cells, and cancer cells. In addition to PDGF, the platelets also contain TGF-β1, HGF, and 5-HT, which are released during the platelet adhesion and aggregation associated with tissue injury [3]. The released PDGF is thought to trigger a series of responses including inflammation and macrophage, neutrophil, and fibroblast migration. RWesearchers investigated the proliferative effects of PDGF on hepatocytes in culture. The results showed that PDGF-BB alone promotes DNA synthesis and proliferation of hepatocytes and that RTK, PI3K, ERK, and mTOR are involved in its signal transduction [16][21]. In addition, an adrenergic α1 receptor agonist enhances the effects of the PDGF-BB, suggesting crosstalk with adrenergic α1 receptor-mediated signaling pathways (Figure 1).Insulin
Human insulin is a heterodimer consisting of an A chain of 21 amino acids and a B chain of 30 amino acids, joined by two disulfide bonds. It is produced by pancreatic B cells and can always act on the liver via the portal bloodstream. The receptor for insulin is a built-in tyrosine kinase, and its activation phosphorylates its intracellular substrate, insulin receptor substrate-1 (IRS-1). Subsequently, signals are transmitted to PI3K and protein kinase B, and glucose transporter-4 is translocated to the cell surface. It has also been reported that blood levels of insulin increase soon after PHx [17][40]. The proliferative effect of insulin on hepatocytes has been said to be a co-mitogen that enhances the action of direct mitogens such as EGF. Therefore, researchwers investigated the effects of insulin in culture. TheOur results showed that insulin alone promotes DNA synthesis and hepatocyte proliferation and that RTK, PI3K, ERK, and mTOR are involved in its signaling [18][19][22,41]. In addition, an adrenergic α1 receptor agonist potentiates the action of insulin, suggesting crosstalk with adrenergic α1 receptor-mediated signaling pathways (Figure 1).2.1.2. Co-Mitogens
Noradrenaline
Noradrenaline is a sympathetic neurotransmitter with a catecholamine structure. Noradrenaline receptors include adrenergic α and β types, each of which has subtypes. All the receptors for noradrenaline are G-protein-coupled ones. It has been reported that sympathetic hyperactivity due to post-PHx invasion increases the blood levels of noradrenaline in about 1 h [20][42]. It activates the duodenal Brunner’s gland and stimulates EGF production and HGF expression [21][22][43,44]. In primary cultured hepatocytes, noradrenaline alone does not promote the proliferation of hepatocytes, but, through crosstalk, it can enhance the hepatocyte proliferation-promoting effects of EGF [8][13], TGF-α [10][19], HGF [14][20], PDGF [16][21], and insulin [19][22] (Figure 1). In addition, in combination with phenylephrine (a selective adrenergic α1-receptor agonist), it enhances the hepatocyte-proliferative effects of EGF, HGF, TGF-α, PDGF, and insulin. Metaproterenol (a selective β2-receptor agonist) enhances the proliferative action of EGF, IGF-I, and HGF. These results show that noradrenaline exhibits unique regulatory mechanisms for these growth factors.2.1.3. Indirect Mitogens
IL-1β
IL-1β was the first inflammatory cytokine among the ILs to be identified, and there are two types, IL-1α and IL-1β [23][45]. IL-1β is a protein consisting of 110–140 amino acids. IL-1 receptors are highly homologous to toll-like receptors and activate serine/threonine kinases via adaptor proteins for intracellular signal transduction. IL-1 is rarely detected in normal tissues and is produced and secreted by macrophages and other immune cells activated by infiltrating inflammation. It has proliferative effects on monocytes and granulocytes. The proliferative effects of IL-1β on hepatocytes have been examined in culture. The results have shown that IL-1β alone promotes DNA synthesis and the proliferation of hepatocytes and that the IL-1β effects are mediated via autocrine secretion of TGF-α from hepatocytes (Figure 2) [24]. IL-1α does not promote the proliferation of hepatocytes.