Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Jarmo-Charles Julian Kalinski and Version 2 by Rita Xu.
Pyrroloiminoquinones are a group of cytotoxic alkaloids most commonly isolated from marine sponges. Structurally, they are based on a tricyclic pyrrolo[4,3,2-de]quinoline core and encompass marine natural products such as makaluvamines, tsitsikammamines and discorhabdins.
- makaluvamine
- damirone
- discorhabdin
- batzelline
- tsitsikammamine
1. Introduction
Pyrroloiminoquinones are a large and diverse group of natural products that have been isolated predominantly from marine sponges [1][2][3][4][1,2,3,4]. They are considered to be potential drug leads due to their significant inhibition of cell proliferation in various cancer cell lines, including promising in vivo activity against several tumor types [5][6][7][8][9][5,6,7,8,9] and inhibition of Plasmodium berghei parasitemia [10] in mouse models. In addition, pyrroloiminoquinones have been shown to exhibit antiviral [11][12][11,12], antifungal [13][14][15][13,14,15] and antibacterial [5][12][15][16][17][5,12,15,16,17] as well as neuromodulatory [18][19][18,19] and antioxidant [20] activities. The mechanisms of bioactivity for these compounds are not yet completely understood with members of this compound class appearing to exert their activity through a number of different modes of action which, for anticancer cell activity, include direct DNA damage [7][8][7,8] and inhibition of key cell regulatory enzymes [21].
Pyrroloiminoquinone molecular structures are characterized by a condensed tricyclic pyrrolo[4,3,2-de]quinoline core that is also considered the principal pharmacophore of this compound class responsible for their antiproliferative and cytotoxic effects [7][22][7,22]. Most compounds of this class can be assigned to one of three major classes exhibiting distinct core structures, namely makaluvamines, bispyrroloiminoquinones and discorhabdins (Figure 1).
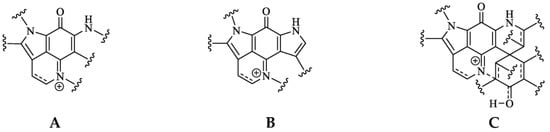
Figure 1. Pyrroloiminoquinone structural scaffolds (A)—Makaluvamines, (B)—Bispyrroloiminoquinones, (C)—Discorhabdins.
Pyrroloiminoquinones have been mostly isolated from marine sponges of the order Poecilosclerida, with Latrunculiidae species from temperate and cold-water environments such as New Zealand, South Africa, the Arctic, and Antarctic as well as warm-water Acarnidae species from the Indo-Pacific proving particularly productive sources [2][3][4][2,3,4]. Nevertheless, members of this class of alkaloids have been reported from ascidians [23][24][23,24] and simple representatives have also been isolated from cultured myxomycetes [25][26][25,26]. Moreover, closely related alkaloids have been reported in hydroids [27][28][27,28], terrestrial fungi [29][30][31][32][29,30,31,32] and marine actinobacteria [33][34][33,34]. This wide geographical and phylogenetic distribution of pyrroloiminoquinone producers, as well as the production of related compounds by bacteria raises the question of microbial involvement in their biosynthesis within marine invertebrates.
2. Structures and Host Distribution of Natural Pyrroloiminoquinones and Related Compounds
2.1. Makaluvamines, Bispyrroloiminoquinones and Discorhabdins
Structurally, the simplest pyrroloiminoquinones are represented by the makaluvamines consisting of the characteristic pyrrolo[4,3,2-de]quinoline core and variable substituents (Figure 2). These include N-methylation of the pyrrole or imine nitrogen, halogenation at C-6, Δ3,4-desaturation and alkylation of N-7 with phenylethyl based side chains. Notable exceptions are makaluvamine O and makaluvamine W and the broad use of the term ‘makaluvamine’ in this review excludes these two structures. Makaluvamines were first reported in 1993 in the sponge Zyzzya fuliginosa collected near the Makaluva Islands, Fiji [7] and since then, have most routinely been isolated from Pacific and Indo-Pacific warm-water sponges of the genus Zyzzya (family Acarnidae) [8][10][11][20][35][36][37][38][39][40][41][42][43][44][8,10,11,20,38,39,40,41,42,43,44,45,46,47]. Makaluvamines have also been isolated from latrunculid sponge species collected off South Africa [22][45][46][22,48,49], the Korean peninsula [16], New Zealand [47][50] and Australia [21]. Interestingly, simple makaluvamines bearing either no substituents or only exhibiting N-methylation have been purified from cultured myxomycetes, Didymium iridis and Didymium bahiense, isolated from Japanese forest litter samples [25][26][25,26]. Makaluvamines are thought to be the biosynthetic precursors to more complex pyrroloiminoquinones and the sulfur-containing makaluvamine F may represent a precursor to sulfur-containing discorhabdins [47][50].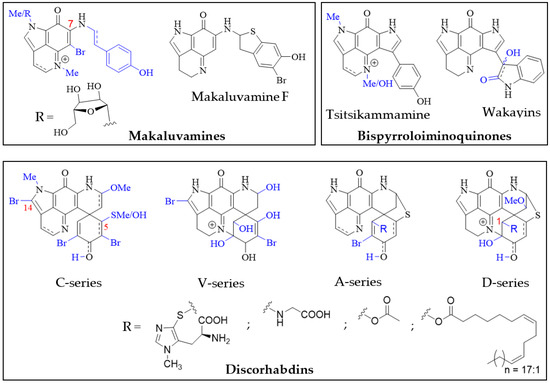
Figure 2. Main pyrroloiminoquinone classes. Variable substituents encompassing all known compound class members are shown in blue.
2.2. Unusual Pyrroloiminoquinones and Related Pyrroloquinolines from Marine Sponges
In addition to the monomeric discorhabdins discussed above, several dimeric and trimeric discorhabdins have been isolated from sponges of the genus Latrunculia (Figure 3). These comprise W-series discorhabdins characterized by a disulfide bridge linking two discorhabdin monomers [64][66][64,66], thioether-linked discorhabdin dimers [70][74][70,74], a C-N-linked discorhabdin C dimer [73], a C-N-linked discorhabdin B dimer as well as a discorhabdin trimer [75]. The saturated discorhabdin W dimer and its monomers have been shown to be interconvertible through reductive cleavage and subsequent UV-irradiation [64], whereas thioether-linked discorhabdin dimers were first discovered as a major degradation product of monomeric discorhabdin stored at −20 °C for a fortnight [70]. Such non-enzymatic dimerization together with the observations that discorhabdins have been shown to be prone to nucleophilic attack at C-1 [70][73][70,73] suggest that at least sulfur-bridged discorhabdin dimers may be generated non-enzymatically in situ [73]. Furthermore, LC-MS/MS-driven molecular networking has provided evidence for numerous discorhabdin di- and trimers, some even incorporating makaluvamines, alongside monomeric A- and D-series discorhabdins in extracts of subantarctic Latrunculia apicalis and South African Cyclacanthia bellae [49][52].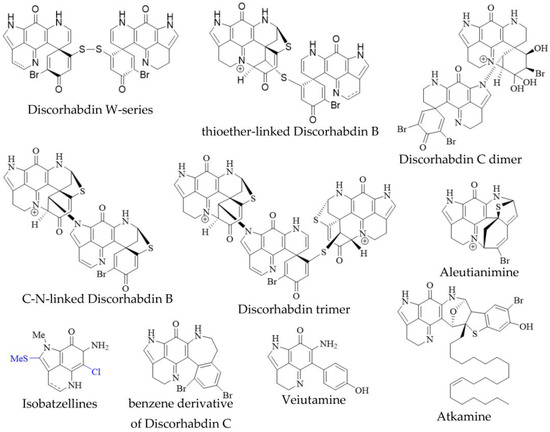
Figure 3. Unusual pyrroloiminoquinones from marine sponges. Variable substituents are shown in blue.
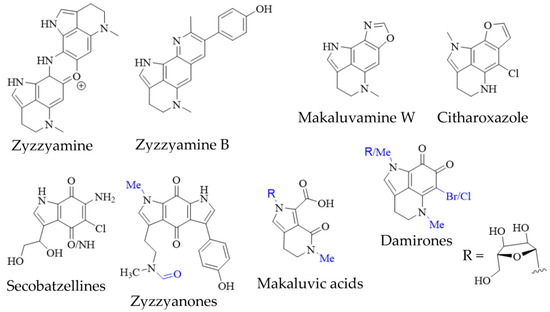
Figure 4. Pyrroloiminoquinone-related marine natural products from sponges. Variable substituents are shown in blue.
2.3. Pyrroloiminoquinones and Related Pyrroloquinolines from Hydroids, Bacteria and Fungi
Pyrroloiminoquinone-related compounds have been isolated from several organisms unrelated to marine sponges or ascidians and this occurrence may hold important information to aid the identification of biosynthetic gene clusters or possibly even microbial symbionts responsible for or involved in pyrroloiminoquinone biosynthesis. Such organisms include the Australian marine hydroid Macrorynchia philippina which contains several cytotoxic macrophilones [27][28][27,28]. Some of these, exhibit a fully formed pyrroloiminoquinone core and have been shown to inhibit the conjugation of SUMO peptides to target proteins, eliciting greatly decreased levels of proteins involved in ERK signaling, while also exhibiting selective cytotoxicity in the NCI-60 anticancer panel [27][28][27,28]. In addition, related pyrroloquinoline alkaloids such as the lymphocyte kinase-inhibiting lymphostin [33] and the selectively cytotoxic ammosamides [34] are produced by marine-derived actinomycetes, Salinispora sp. and Streptomyces sp., respectively (Figure 5).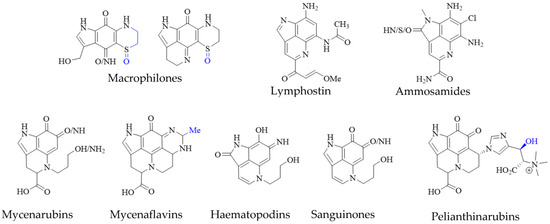
Figure 5. Pyrroloiminoquinone-related natural products from marine hydroids, marine soil-derived bacteria, and terrestrial fungi. Variable substituents are shown in blue.