Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Nicole C. Riedel and Version 2 by Camila Xu.
Malignant brain neoplasms are a heterogeneous group of tumors, including glioma, ependymoma, embryonal tumors, and many other (rare) entities and subentities, affecting patients from birth to adulthood. Organoids emerged as three-dimensional (3D) cell culture systems for modeling healthy and diseased tissues. These organoids potentially model development, diseases, and drug responses [13]. They are self-organizing three-dimensional structures that closely mimic an organ or tissue at a morphological, cellular, and functional level.
- organoids
- precision medicine
- pediatric
- Medulloblastoma
- pediatric glioma
- ETMR
- ATRT
- pediatric brain tumors
1. Introduction
Malignant brain neoplasms are a heterogeneous group of tumors, including glioma, ependymoma, embryonal tumors, and many other (rare) entities and subentities, affecting patients from birth to adulthood. Despite intensive treatment protocols, including surgery, chemotherapy, and radiotherapy, the prognosis for many high-grade brain tumor patients remains poor [1][2][3][4][5][6][1,2,3,4,5,6]. Although extensive research in this field has resulted in a detailed molecular classification of brain tumors and led to various new insights into their biology, numerous recent clinical trials have failed to significantly improve the prognosis for these patients, especially those suffering a relapse [7]. Overall, there is a noticeable gap between recent preclinical achievements and the clinical improvements in patient outcomes. This dichotomy may stem from current preclinical studies frequently being conducted in two-dimensional (2D) cell culture, which neither sufficiently recapitulates inter- nor intratumoral heterogeneity nor the cellular diversity of the tumor microenvironment (TME) [8]. Importantly, heterogeneous in vivo-like tumor cell populations respond differently to drug treatment than 2D in vitro entities, and intratumoral diversity results in a higher risk of treatment resistance and tumor recurrence [9][10][9,10]. Furthermore, the interplay of tumor cells with their TME, including endothelial cells, immune cells, and neuronal cells, affects the treatment response [11][12][11,12]. Therefore, it has become paramount to establish models which include these key characteristics and allow tumor–TME interactions, thus potentially generating more accurate predictions of tumor biology, drug efficacy, and immune response.
In the past few years, organoids emerged as three-dimensional (3D) cell culture systems for modeling healthy and diseased tissues. These organoids potentially model development, diseases, and drug responses [13]. They are self-organizing three-dimensional structures that closely mimic an organ or tissue at a morphological, cellular, and functional level. They can be divided into two major groups: organoids that mirror healthy tissue, including brain organoids, and those that simulate diseased tissue, including tumor organoids [13]. Initial organoid models utilized the intrinsic self-patterning abilities of human pluripotent stem cells in appropriate conditions to generate small aggregates with optic cups [14] or even tissues representing a wide gamut of brain regions, forming the so-called cerebral organoids [15]. Later on, other groups investigated organoids resembling specific brain regions, including the forebrain, the midbrain, the hypothalamus, or the cerebellum [16][17][18][16,17,18]. Pioneering the use of organoids for tumor research, Sato et al. generated 3D in vitro models from primary colon carcinoma samples [19]. They were followed by other groups who developed tumor organoids from various entities, including prostatic, pancreatic, and liver cancers, as well as glioblastoma [20][21][22][23][20,21,22,23].
These tumor organoids can then be used for high-throughput drug and toxicity screenings to uncover new personalized therapeutics [13][24][25][26][13,24,25,26]. Additionally, healthy tissue 3D cell culture systems, such as organoids or organ-on-a-chip models, can be further used to test the side-effects of drugs. These technologies may identify drugs with a high efficacy against the tumor and a low burden of side effects on healthy tissues [24][27][28][24,27,28]. Recapitulating parental tumors, cancer organoids have proven to be capable of predicting cancer treatment efficiency in vivo [23][29][30][23,29,30], thus ringing in the era of organoid-based in vitro anti-cancer drug tests.
2. Organoids Are Superior to Prior 2D In Vitro Models in Recapitulating the Primary Tumor Characteristics
Despite making progress on exploring the mechanisms leading to tumor initiation by identifying, for example, (I) the mutational burden of tumors, (II) malignancies’ cells of origin, and (III) the impact of the TME on tumor cells [31][32][33][34][31,32,33,34], many key scientific questions to finally improve brain tumor patients’ survival remain unanswered. This lack of fundamental insight may partly stem from in vitro models insufficiently recapitulating the core characteristics of the primary tumors. For decades, brain tumor research has been based on 2D and 3D cell cultures in mono-layers and spheroids, respectively. Traditional 2D in vitro cultures of tumor models rely on cell propagation in standard petri dishes. These 2D cell culture models undergo clonal selection for fast-growing and cell-culture-compatible cell populations, thereby losing cellular diversity and often resulting in a homogeneous cell population that are no longer recapitulating the tumors’ original heterogeneity [8][35][36][8,35,36]. Due to their mono-layer arrangement, these cell cultures are adapted to conditions of 20% oxygen, which exceeds the usual oxygen level of about 5% in in vivo tumors [8][35][8,35]. Spheroids consist of mostly uniform aggregates of a mixture of desired and relevant cell types for a given disease model assembled in an essentially random three-dimensional arrangement. On the other hand, organoids self-arrange their cell types into micro-moieties that more closely approximate organ tissue structure and function. With the advent of these self-organizing organoid tissues, a plethora of more complex three-dimensional model systems have addressed brain tumors in recent years, including patient-derived tumor organoids (PDO), patient-derived explants (PDEs), tumor-brain organoids (TBOs), neoplastic cerebral organoids (neoCORs), and, lastly, bioprinted tumor models (Figure 1). These organoid models can be divided into two major groups: (I) tumoroids starting only from a tumor tissue, including PDOs and PDEs, and (II) organoids composed of a tumor and a non-tumor compartment, including TBOs and neoCORs.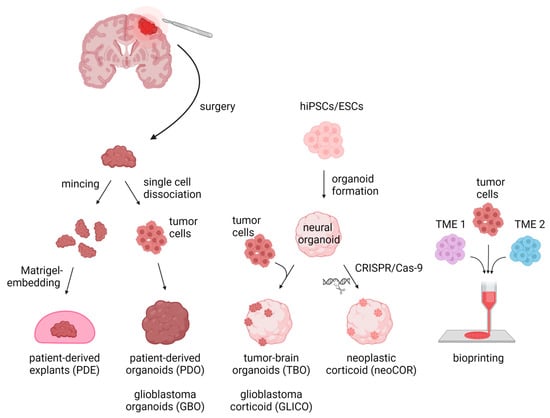
Figure 1. Three-dimensional in vitro tumor models can be derived from either primary patient materials or generated de novo from pluripotent stem cells. PDEs and PDOs are derived from patients’ resected tumor tissue, which is minced and Matrigel-embedded for PDE generation or is single-cell dissociated followed by re-aggregation for PDOs/GBOs. TBOs/GLICOs and neoCORs are generated by seeding hiPSCs/ESCs for organoid generation, followed by co-culture with tumor cells for TBOs/GLICOs or CRISPR-Cas9-based gene editing for neoCORs. To create 3D models via bioprinting, tumor cells are loaded together with TME cells into a bioink and spatially printed. Abbreviations: PDE: patient-derived explant; PDO: patient-derived tumor organoid; GBO; glioblastoma organoid; TBO: tumor-brain organoid; GLICO: glioblastoma corticoid; neoCOR: neoplastic corticoid; 3D: three-dimensional; hiPSC: human induced pluripotent stem cell; ESC: embryonic stem cell; TME1/2: tumor microenvironment cell type 1 or 2. Created with BioRender.com (accessed on 7 November 2022).
3. Current Limitations of Organoids
While 3D model systems have improved over recent years, they still face several limitations. To properly mimic a tumor in vitro, current organoid models are missing vascularization, TME cells (immune cells and neuronal TME), and protocols resulting in more reproducible and scalable organoid generation (Figure 2).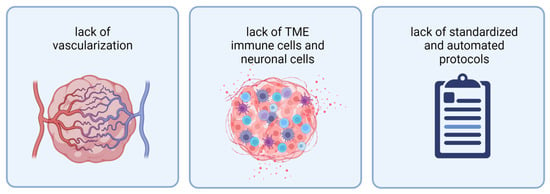
Figure 2. Current organoids face three limitations: from left to right: lack of vascularization, lack of TME immune cells and neuronal cells, and lack of standardized and automated protocols for organoid generation. Abbreviations: TME: tumor microenvironment. Created with BioRender.com (accessed on 7 November 2022).
3. Organoid Models in Pediatric Brain Tumors
Pediatric brain tumors are much rarer than brain tumors in adults but belong to the most frequent tumor entities in children. Besides pediatric glioma and ependymoma, children are affected by embryonal tumors, including medulloblastoma, atypical teratoid and rhabdoid tumor (ATRT), and embryonal tumor with multilayered rosettes (ETMR), most of which are associated with a poor prognosis [59][85]. Despite recent achievements in establishing organoid models for adult brain tumors, such as glioblastoma or LGG [29][40][54][60][29,40,54,84], a similar body of work for pediatric brain tumors is still missing (Table 1). Unfortunately, the results gathered from in vitro tumor models cannot necessarily be transferred from adult to pediatric entities as tumor location and molecular characteristics of the tumor and the developmental state of the brain, as a key player of the TME, differ. Recent studies have shown that drugs that work perfectly in adults may cause major side effects on the developing brain of children [61][86]. Nevertheless, drugs are primarily established for adults and then transferred to pediatric patients [61][86]. Using 3D cell culture techniques might help to overcome this problem and directly establish drugs for this age group. Thus, one major challenge is the development of individual model systems for each pediatric brain tumor entity and subentity. In the last few years, some groups started testing PDO or neoCOR models for some of these entities [52][62][63][64][52,87,88,89]. Due to the scarcity of publications, the next section will summarize recent achievements by entity and not by model.Table 1.
Established three-dimensional brain tumor models.
| Reference | Entity | Model-Type | Method |
|---|---|---|---|
| Hubert et al., 2016 [23] | glioblastoma | GBO | Tumor cells embedded in Matrigel |
| Jacob et al., 2020 [29] | glioblastoma | GBO | Tumor pieces on an orbital shaker |
| Loong et al., 2020 [65][62] | glioblastoma | GBO | Tumor cells embedded in Matrigel |
| Chen et al., 2022 [43] | glioblastoma | GBO | Tumor pieces on an orbital shaker |
| LeBlanc et al., 2022 [37] | glioblastoma | PDE | Tumor pieces in Matrigel |
| da Silva et al., 2018 [66][67] | glioblastoma | GLICO | Murine brain organoids, GBM cells |
| Linkous et al., 2019 [40] | glioblastoma | GLICO | Brain organoids + GBM cells |
| Krieger et al., 2020 [54] | glioblastoma | GLICO | Brain organoids + GBM cells |
| Gorancia-Buzhala et al., 2020 [67][69] | glioblastoma | GLICO | Brain organoids + GBM cells |
| Azzarelli et al., 2021 [68][70] | glioblastoma | GLICO | Brain organoids + GBM cells |
| Ogawa et al., 2018 [69][75] | glioblastoma | neoCOR | HRas, TP53 mutations |
| Bian et al., 2018 [53] | glioblastoma | neoCOR | Several different mutations in combination and alone as PTEN, Myc, and EGFR |
| Yi et al., 2019 [70][78] | glioblastoma | bioprinting | GBM cells + endothelial cells + HUVECs |
| Heinrich et al., 2019 [71][76] | glioblastoma | bioprinting | GBM cells + macrophages |
| Tang et al., 2020 [72][77] | glioblastoma | bioprinting | GBM cells + neuronal progenitor cells + astrocytes + macrophages |
| Abdullah et al., 2022 [60][84] | LGG | PDO | Tumor pieces on an orbital shaker; 5% O2 |
| Sundar et al., 2022 [62][87] | pediatric HGG | PDO | Tumor cells embedded in Matrigel |
| Frisira et al., 2019 [64][89] | medulloblastoma | PDO | Tumor cells embedded in Matrigel |
| Ballabio et al., 2020 [63][88] | medulloblastoma | neoCOR | Different mutations, e.g., Otx-2 or c-Myc |
| Parisian et al., 2020 [52] | ATRT | neoCOR | SMARCB1-KD |