Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 3 by Conner Chen and Version 2 by Conner Chen.
Climate change and the urgent need to reduce greenhouse gas (GHG) emission from agriculture has resulted in significant pressure on the livestock industry for advanced practices that are environmentally more sustainable. Livestock is responsible for more than 15% of anthropogenic methane (CH4) emission via enteric fermentation and improved strategies for mitigating enteric CH4 production therefore represents a promising target to reduce the overall GHG contribution from agriculture. Ruminal CH4 is produced by methanogenic archaea, combining carbon dioxide (CO2) and hydrogen (H2). Removal of H2 is essential, as its accumulation inhibits many biological functions that are essential for maintaining a healthy rumen ecosystem.
- hydrogenotrophy
- methanogenesis
- propionogenesis
- reductive acetogenesis
1. Introduction
The rumen harbors a highly diverse and complex mixture of microorganisms, including archaea (108−109/mL), bacteria (1010−1011/mL), ciliate protozoa (106/mL), and fungi (106/mL), which facilitate the degradation of complex plant carbohydrates into small molecules [1] and ultimately provide metabolites that can be used by the ruminant animal [2][3][4][5]. Livestock are mainly fed with agricultural crops, which via microbial activity are converted to metabolic intermediates (i.e., volatile fatty acids (VFAs), such as acetate, butyrate and propionate, and hydrogen (H2) and gaseous end products such as carbon dioxide (CO2) and methane (CH4) [6]. Increased microbial H2 production and its subsequent accumulation, which can be promoted by a high-starch diet, have several detrimental effects on the rumen ecosystem and that can be attributed to a decrease in rumen pH triggered by starch fermentation. These effects include the deactivation of specific biomass-degrading enzymes from some of the most efficient fiber degraders of the rumen microbiome but also system-level responses, such as the reduction of feed conversion within the rumen [7][8]. Methanogens, a group of microbes belonging to the phylogenetic group of the archaea, combine molecular H2 with CO2 to produce CH4 during methanogenesis, enabling the removal of H2 from the system [9][10]. Although this removal of H2 is important for maintaining a healthy rumen ecosystem, from the viewpoint of nutrient expenditure methanogenesis is a costly process, accounting for a gross energy intake loss of 2–12% in ruminants [11][12][13][14]. Since the annual production of enteric CH4 accounts for ~15% of total anthropogenic CH4 emissions [11][15], with CH4 having a global warming potential 23-fold higher than that of CO2, there is also a real and severe environmental cost associated with the energy of the enteric CH4 that is released into the atmosphere.
Strategies and factors for CH4 abatement have been reviewed in the past [1][9][12][16][17][18][19][20][21][22][23][24][25] and many of the strategies used to mitigate CH4 from ruminants involve the use of antibiotics, ionophores [26], halogenated CH4 analogues [27][28][29], heavy metals [30], lipid-rich materials such as coconut oil [31][32][33], probiotics [27], bacteriocin [34], and numerous chemicals [35][36]. Immunization against methanogens [37][38], elimination of ciliate protozoa (defaunation) both in in vivo and in vitro [39] and addition of acetogenic bacteria to rumen fluid [40][41][42] in in vitro experiments have also been tested. Use of toxic chemicals and antibiotics as inhibitors, although considered an option in the past, are no longer accepted due to rising concerns regarding their impact on the environment, the animal, and potentially on the consumer of the animal products [43]. Interventions using phage therapy, altering methanogenic diversity and chemogenomic approaches [6] are some of the more recent technologies, but the extent to which these processes remove and eliminate the produced H2 still remains to be investigated. Therefore, a critical step for a successful CH4 reduction strategy may be one that uses natural processes within the rumen. One such approach relies on establishing a non-methanogenic sink for H2 produced during fermentation.
2. Hydrogen: A Key Player in Rumen Fermentation
H2 concentration plays a major role in the regulation of microbial fermentation in the rumen [44][45][46][47]. The partial H2 pressure is a key regulator of H2 metabolism and the fate of ruminal H2 disposal with dissolved H2 gas and H2 ion determining the redox potential of the rumen liquor. The efficient elimination of H2 enhances fermentation by reducing its inhibitory effect on microbial growth and microbial degradation of plant material [48][49]. Destiny of H2 liberation is associated with favorable thermodynamic changes and an inverse correlation between Gibbs free energy (ΔG0) and the minimum partial H2-pressure that is required for a reaction to continue: a reaction is considered to be thermodynamically more competitive when its requirement of H2 partial pressure is low [50]. Due to this central regulatory role in rumen fermentation, H2 can be considered to be the currency of ruminal fermentation [51]. Removal of the major fraction of the rumen H2 occurs via the methanogenic archaea to CH4, during which four moles of H2 are consumed and converted into one mole of CH4, which is then released into the atmosphere though eructation. During hydrogenotrophic methanogenesis, methanogens use CO2 as carbon source and terminal electron acceptor and H2 as electron donor. Other non-methanogenic rumen microbes, using CO2 and other electron acceptors such as sulfate, nitrate, and fumarate, compete with methanogens for H2, but they play a less dominant role in the removal of H2 from the rumen ecosystem [52][53]. Non-methanogenic bacteria that use H2 as electron donor include acetogens that reduce CO2 to form acetate by the Wood-Ljungdahl pathway [54], sulfate-reducing bacteria (SRB) that reduce sulfate to hydrogen sulfide [55], nitrate-reducing bacteria (NRB) that reduce nitrate (NO3) to ammonia (NH4) and fumarate-reducing bacteria that use H2 to form succinate [56][57]. Succinate can subsequentially be decarboxylated to propionate, which is a valuable nutrient for the ruminant animal [58], either by the succinate producer itself or it can be transferred to succinate users as an intercellular electron carrier [45]. Figure 1 summarizes the microbial pathways for H2 removal from the rumen.
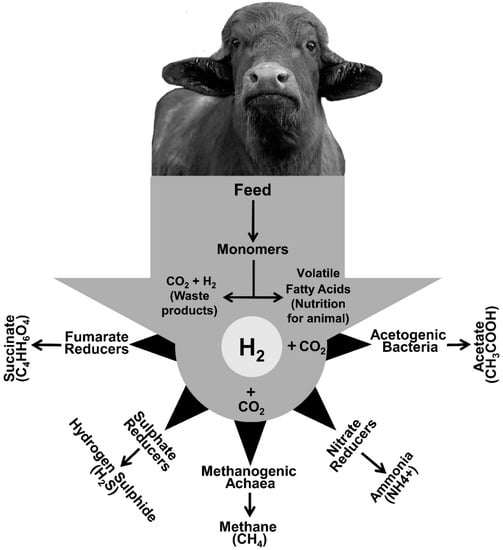
Figure 1. Major and minor H2 and CO2 sequestering pathways in rumen.
Ruminal methanogenesis, contributing CH4 to the atmosphere, is directly and inversely linked to the animal productivity. The ability to control CH4 emission especially reduce methanogenesis from agriculture has enormous environmental and socioeconomic implication, but it also requires a detailed understanding of the microorganisms and microbial processes that are involved. Although a complete understanding of these highly interwoven microbial and metabolic networks has still not been achieved and most likely will not be feasible in the immediate future, there are some aspects that are reasonably well understood. These aspects represent a promising starting point for targeted CH4 reduction from ruminants. One of the promising key intermediates that has been recognized as such and that has received significant attention for targeted CH4 mitigation is metabolic H2 and the metabolic pathways, microbes and enzymes involved its production and consumption.
Since H2 is an immediate precursor for the archaeal reduction of CO2 into CH4, biological approaches that redirect H2 away from archaeal methanogenesis and into alternate metabolic pathways seem to be the most promising approaches to convert feed carbon into metabolic energy for the ruminant instead of releasing it into the atmosphere. Redirecting H2 through reductive acetogenesis and propionogenesis has advantages over other pathways due to production of valuable metabolic end products that can be used by the host animal as nutrients and can be converted into animal proteins for human consumption. Although the understanding of how to redirect metabolic H2 into more favorable pathways facilitates the production of value-added metabolic intermediates and therefore redirects otherwise lost feed energy, several issues related to the fine tuning of this redirection, such as the co-factor requirements, toxicity of metabolic intermediates, as well as thermodynamics of competing metabolic processes, need to be investigated in greater detail. A further aspect that will have to be investigated in future and that will have direct implications for the translational value of findings on the area of rumen nutrition and function is the link and dependence of the rumen microbiome and its function in dietary conversion.
References
- Wang, S.; Giller, K.; Kreuzer, M.; Ulbrich, S.E.; Braun, U.; Schwarm, A. Contribution of ruminal fungi, archaea, protozoa, and bacteria to the methane suppression caused by oilseed supplemented diets. Front. Microbiol. 2017, 8, 1864.
- Choudhury, P.K.; Salem, A.Z.M.; Jena, R.; Kumar, S.; Singh, R.; Puniya, A.K. Rumen microbiology: An overview. In Rumen Microbiology-Evolution to Revolution; Puniya, A.K., Singh, R., Kamra, D.N., Eds.; CRC Springer: New Delhi, India, 2015; pp. 3–16.
- Kumar, S.; Dagar, S.S.; Puniya, A.K.; Upadhyay, R.C. Changes in methane emission, rumen fermentation in response to diet and microbial interactions. Res. Vet. Sci. 2013, 94, 263–268.
- Kamra, D.N. Rumen microbial ecosystem. Curr. Sci. 2005, 89, 124–135.
- Janssen, P.H.; Kirs, M. Structure of the archaeal community of the rumen. Appl. Environ. Microbiol. 2008, 74, 3619–3625.
- Kumar, S.; Choudhury, P.K.; Carro, M.D.; Griffith, G.W.; Dagar, S.S.; Puniya, M.; Calabro, S.; Ravella, S.R.; Dhewa, T.; Upadhyay, R.C.; et al. New aspects and strategies for methane mitigation from ruminants. Appl. Microbiol. Biotechnol. 2014, 98, 31–44.
- Ragsdale, S.W.; Pierce, E. Acetogenesis and the Wood-Ljungdahl pathway of CO(2) fixation. Biochim. Biophys. Acta 2008, 1784, 1873–1898.
- Ishaq, S.L.; AlZahal, O.; Walker, N.; McBride, B. An investigation into rumen fungal and protozoal diversity in three rumen fractions, during high-fiber or grain-induced sub-acute ruminal acidosis conditions, with or without active dry yeast supplementation. Front. Microbiol. 2017, 8, 1943.
- Malik, P.K.; Bhatta, R.; Gagen, E.J.; Sejian, V.; Soren, N.M.; Prasad, C.S. Alternate H2 sinks for reducing rumen methanogenesis. In Climate Change Impact on Livestock: Adaptation and Mitigation; Sejian, V., Gaughan, J., Baumgard, L., Prasad, C., Eds.; Springer: New Delhi, India, 2015; pp. 303–320.
- Joblin, K. Methanogenic archaea. In Methods in Gut Microbial Ecology for Ruminants; Makker, H., McSweeney, C.S., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 47–53.
- Moss, A.R.; Jouany, J.P.; Newbold, J. Methane production by ruminants: Its contribution to global warming. Ann. Zootech. 2000, 49, 231–253.
- Ungerfeld, E.M. Inhibition of rumen methanogenesis and ruminant productivity: A meta-analysis. Front. Vet. Sci. 2018, 5, 113.
- Beauchemin, K.A.; Ungerfeld, E.M.; Eckard, R.J.; Wang, M. Review: Fifty years of research on rumen methanogenesis: Lessons learned and future challenges for mitigation. Animal 2020, 14, s2–s16.
- Van Nevel, C.J.; Demeyer, D.I. Control of rumen methanogenesis. Environ. Monit. Assess. 1996, 42, 73–97.
- Demeyer, D.; Fiedler, D.; De Graeve, K.G. Attempted induction of acetogenesis into the rumen fermentation in vitro. Reprod. Nutr. Dev. 1996, 36, 233–240.
- Garsa, A.K.; Choudhury, P.K.; Puniya, A.K.; Dhewa, T.; Malik, R.K.; Tomar, S.K. Bovicins: The bacteriocins of streptococci and their potential in methane mitigation. Probiotics Antimicrob. Proteins 2019, 11, 1403–1413.
- Goopy, J.P. Creating a low enteric methane emission ruminant: What is the evidence of success to the present and prospects for developing economies? Anim. Prod. Sci. 2019, 59, 1769–1776.
- Hook, S.E.; Wright, A.D.G.; McBride, B.W. Methanogens: Methane producers of the rumen and mitigation strategies. Archaea 2010, 2010, 945785.
- Kim, S.H.; Mamuad, L.L.; Islam, M.; Lee, S.S. Reductive acetogens isolated from ruminants and their effect on in vitro methane mitigation and milk performance in Holstein cows. J. Anim. Sci. Technol. 2020, 62, 1–13.
- Martin, C.; Morgavi, D.P.; Doreau, M. Methane mitigation in ruminants: From microbe to the farm scale. Animal 2010, 4, 351–365.
- Wanapat, M.; Kongmun, P.; Poungchompu, O.; Cherdthong, A.; Khejornsart, P.; Pilajun, R.; Kaenpakdee, S. Effects of plants containing secondary compounds and plant oils on rumen fermentation and ecology. Trop. Anim. Health Prod. 2012, 44, 399–405.
- Beauchemin, K.A.; Kreuzer, M.O.; O’Mara, F.P.; McAllister, T.A. Nutritional management for enteric methane abatement: A review. Aust. J. Exp. Agric. 2008, 48, 21–27.
- García-González, R.; López, S.; Fernández, M.; Bodas, R.; González, J.S. Screening the activity of plants and spices for decreasing ruminal methane production in vitro. Anim. Feed Sci. Technol. 2008, 147, 36–52.
- Eckard, R.J.; Grainger, C.; de Klein, C.A.M. Options for the abatement of methane and nitrous oxide from ruminant production: A review. Livest. Sci. 2011, 130, 47–56.
- Patra, A.; Park, T.; Kim, M.; Yu, Z. Rumen methanogens and mitigation of methane emission by anti-methanogenic compounds and substances. J. Anim. Sci. Biotechnol. 2017, 8, 13.
- McGinn, S.M.; Beauchemin, K.A.; Coates, T.; Colombatto, D. Methane emissions from beef cattle: Effects of monensin, sunflower oil, enzymes, yeast and furmaric acid. J. Anim. Sci. 2004, 82, 3346–3356.
- Janssen, P.H.; Frenzel, P. Inhibition of methanogenesis by methyl fluoride: Studies of pure and defined mixed cultures of anaerobic bacteria and archaea. Appl. Environ. Microbiol. 1997, 63, 4552–4557.
- Miller, T.L.; Wolin, M.J. Inhibition of growth of methane-producing bacteria of the ruminant forestomach by hydroxymethylglutaryl-SCoA reductase inhibitors. J. Dairy Sci. 2001, 84, 1445–1448.
- Mitsumori, M.; Shinkai, T.; Takenaka, A.; Enishi, O.; Higuchi, K.; Kobayashi, Y.; Nonaka, I.; Asanuma, N.; Denman, S.E.; McSweeney, C.S. Responses in digestion, rumen fermentation and microbial populations to inhibition of methane formation by a halogenated methane analogue. Br. J. Nutr. 2012, 108, 482–491.
- Sanchez, J.M.; Valle, L.; Rodriguez, F.; Morinnigo, M.A.; Borrego, J.J. Inhibition of methanogenesis by several heavy metals using pure cultures. Lett. Appl. Microbiol. 1996, 23, 439–444.
- Fievez, V.F.; Dohme, M.; Daneels, K.R.; Demeyer, D. Fish oils as potent rumen methane inhibitors and associated effects on rumen fermentation in vitro and in vivo. Anim. Feed Sci. Technol. 2003, 104, 41–58.
- Machmüller, A.; Kreuzer, M. Methane suppression by coconut oil and associated effects on nutrient and energy balance in sheep. Can. J. Anim. Sci. 1999, 79, 65–72.
- Machmüller, A. Medium-chain fatty acids and their potential to reduce methanogenesis in domestic ruminants. Agric. Ecosyst. Environ. 2006, 112, 107–114.
- Lee, S.S.; Hsu, J.T.; Mantovani, H.C.; Russell, J.B. The effect of bovicin HC5, a bacteriocin from Streptococcus bovis HC5, on ruminal methane production in vitro. FEMS Microbiol. Lett. 2002, 217, 51–55.
- Baker, S.K. Rumen methanogens and inhibition of methanogenesis. Aust. J. Agric. Res. 1999, 50, 1293–1298.
- Anderson, R.C.; Carstens, G.E.; Miller, R.K.; Callaway, T.R.; Schultz, C.L.; Edrington, T.S.; Harvey, R.; Nisbet, D. Effect of oral nitroethane and 2-nitropropanol administration on methane-producing activity and volatile fatty acid production in the ovine rumen. Bioresour. Technol. 2006, 97, 2421–2426.
- Wright, A.D.G.; Kennedy, P.; O’neill, C.J.; Toovey, A.F.; Popovski, S.; Rea, S.M.; Pimm, C.L.; Klein, L. Reducing methane emissions in sheep by immunization against rumen methanogens. Vaccine 2004, 22, 3976–3985.
- Zhang, L.; Huang, X.; Xue, B.; Peng, Q.; Wang, Z.; Yan, T.; Wang, L. Immunization against rumen methanogenesis by vaccination with a new recombinant protein. PLoS ONE 2015, 10, e0140086.
- Newbold, C.J.; de la Fuente, G.; Belanche, A.; Ramos-Morales, E.; McEwan, N.R. The Role of Ciliate Protozoa in the Rumen. Front. Microbiol. 2015, 6, 1313.
- LeVan, T.D.; Robinson, J.A.; John, R.; Greening, R.C.; Smolenski, W.J.; Leedle, J.A.Z.; Schaefer, D.M. Assessment of reductive acetogenesis with indigenous ruminal bacterium populations and Acetitomaculum ruminis. Appl. Environ. Microbiol. 1998, 64, 3429–3436.
- Nollet, L.; Demeyer, D.; Verstraete, W. Effect of 2-bromoethanesulfonic acid and Peptostreptococcus productus ATCC35244 addition on stimulation of reductive acetogenesis in the ruminal ecosystem by selective inhibition of methanogenesis. Appl. Environ. Microbiol. 1997, 63, 194–200.
- Nollet, L.; Velde, I.V.; Verstraete, W. Effect of the addition of Peptostreptococcus productus ATCC35244 on the gastro-intestinal microbiota and its activity, as simulated in an in vitro simulator of the human gastro-intestinal tract. Appl. Microbiol. Biotechnol. 1997, 48, 99–104.
- Liu, H.; Wang, J.; Wang, A.; Chen, J. Chemical inhibitors of methanogenesis and putative applications. Appl. Microbiol. Biotechnol. 2011, 89, 1333–1340.
- Hungate, R.E. The Rumen and Its Microbes; Academic Press: New York, NY, USA, 1966; p. 533.
- Ungerfeld, E.M. Metabolic hydrogen flows in rumen fermentation: Principles and possibilities of interventions. Front. Microbiol. 2020, 11, 589.
- Williams, A.G.; Coleman, G.S. The rumen protozoa. In The Rumen Microbial Ecosystem, 2nd ed.; Hobson, P.N., Stewarteds, C.S., Eds.; Blackie Academic & Professional: New York, NY, USA, 1997; pp. 73–139.
- Cord-Ruwisch, R.; Seitz, H.J.; Conrad, R. The capacity of hydrogenotrophic anaerobic bacteria to compete for traces of hydrogen depends on the redox potential of the electron acceptor. Arch. Microbiol. 1988, 149, 350–357.
- Immig, I. The rumen and hindgut as a source of ruminant methanogenesis. Environ. Monit. Assess. 1996, 42, 57–72.
- McAllister, T.A.; Newbold, C.J. Redirecting rumen fermentation to reduce methanogenesis. Aust. J. Exp. Agric. 2008, 48, 7–13.
- Ellis, J.L.; Dijkstra, J.; Kebreab, E.; Bannink, A.; Odongo, N.E.; Mcbride, B.W.; France, J. Aspects of rumen microbiology central to mechanistic modelling of methane production in cattle. J. Agric. Sci. 2008, 146, 213–233.
- Hegarty, R.S.; Gerdes, R. Hydrogen production and transfer in the rumen. Rec. Adv. Anim. Nutr. 1999, 12, 37–44. Available online: http://livestocklibrary.com.au/handle/1234/19891 (accessed on 25 October 2022).
- Greening, C.; Geier, R.; Wang, C.; Woods, L.C.; Morales, S.E.; McDonald, M.J.; Rushton-Green, R.; Morgan, X.C.; Koike, S.; Leahy, S.C.; et al. Diverse hydrogen production and consumption pathways influence methane production in ruminants. ISME J. 2019, 13, 2617–2632.
- Morgavi, D.P.; Forano, E.; Martin, C.; Newbold, C.J. Microbial ecosystem and methanogenesis in ruminants. Animal 2010, 4, 1024–1036.
- Gagen, E.J.; Denman, S.E.; McSweeney, C.S. Acetogenesis as an alternative to methanogenesis in the rumen. In Livestock Production and Climate Change; Malik, P.K., Bhatta, R., Takahashi, J., Kohn, R.A., Prasad, C.S., Eds.; CABI: Wallingford, UK, 2015; pp. 292–303.
- Morvan, B.; Bonnemoy, F.; Fonty, G.; Gouet, P. Quantitative determination of H2-utilizing acetogenic and sulfate-reducing bacteria and methanogenic archaea from digestive tract of different mammals. Curr. Microbiol. 1996, 32, 129–133.
- Kim, S.H.; Mamuad, L.L.; Kim, D.W.; Kim, S.K.; Lee, S.S. Fumarate reductase-producing enterococci reduce methane production in rumen fermentation in vitro. J. Microbiol. Biotechnol. 2016, 26, 558–566.
- Mamuad, L.L.; Kim, S.H.; Lee, S.S.; Cho, K.K.; Jeon, C.O.; Lee, S.S. Characterization, metabolites and gas formation of fumarate reducing bacteria isolated from Korean native goat (Capra hircus coreanae). J. Microbiol. 2012, 50, 925–931.
- Wolin, M.J.; Miller, T.L.; Stewart, C.S. Microbe-microbe interactions. In The Rumen Microbial Ecosystem; Hobson, P.N., Stewart, C.S., Eds.; Chapman and Hall: London, UK, 1997; pp. 467–488.
More