2. The Peculiar Properties of Human mitoNEET: A Unique Folding for a Multiplicity of Functions
The outer mitochondrial membrane (OMM) protein mitoNEET, also known as CDGSH Fe-S domain-containing protein-1 (CISD1), is composed of 108 amino acids, encompassing a N-terminal transmembrane helix (residues 14–32) that anchors the protein to the OMM
[11], and a cytosolic portion (residues 33–108) that has been widely investigated through X-ray crystallography, showing a unique, highly conserved folding
[12][13][14][15][12,13,14,15]. All the crystallographic structures of mitoNEET revealed the presence of two distinct domains: a β-rich or “β-cap” region and a cluster-binding domain (
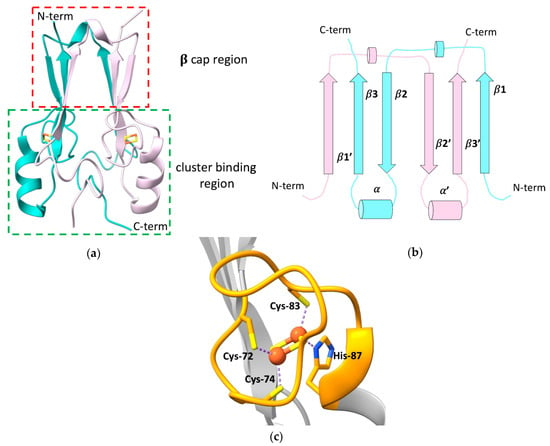
Figure 1a)
[12][13][15][16][12,13,15,16]. The latter contains the highly conserved CXCX
2(S/T)X
3PXCDG(S/A/T)H motif that binds a [2Fe-2S] cluster, and that is part of the characteristic CDGSH-type domain of 39 amino acids, which constitutes the hallmark of the “NEET” family.
Figure 1. (a) Crystallographic structure of the soluble domain of human mitoNEET, with highlighted the β-cap and the cluster-binding regions (PDB ID: 2QH7). The two subunits forming the dimer are reported in different colors; (b) topology diagram illustrating the organization of the secondary structural units in the two protomers of the dimeric structure of mitoNEET; (c) [2Fe-2S] cluster-binding motif of human mitoNEET. The conserved CXCX2(S/T)X3PXCDG(S/A/T)H motif is highlighted in orange.
mitoNEET forms an intertwined, parallel homodimer, with a pseudo two-fold symmetry
[12][13][16][12,13,16]. Each protomer of the dimer comprises a long, flexible loop in the N-terminal part (res 33–55), whose structure has not been solved yet
[14], and displays a β1-β2-α-β4 topology in the rest of the protein (
Figure 1b).
In the dimer, the β-strands form two symmetric β-sheets, composed of two antiparallel β-strands from one monomer (residues 68–71 and 101–104) and an additional parallel swapped-strand from the other monomer (residues 56–61)
[12][13][15][12,13,15]. These two β-sheets and the two loops connecting the swapped β-strands and also containing a helical turn (residues 62–64), form the so-called β-cap region
[12][13][15][16][12,13,15,16], whereas the N-terminal portion of the soluble domain, the conserved α-helix formed by residues 86–94 and the loop connecting the helix to the β-cap, together with the CXCX
2(S/T)X
3PXCDG(S/A/T)H motif, form the cluster-binding domain
[12][13][15][16][12,13,15,16]. Here, two symmetric hydrophobic cores, comprising Ile-45, Ile-56, Trp-75, Phe-80 of one monomer, and Val-98 of the second monomer
[13], and two intermolecular hydrogen bonds between His-58 and Arg-73
[15], further stabilize the dimeric state of mitoNEET.
The folding of the two domains is indeed interdependent. Theoretical structure-based folding studies proposed that the rigidity in the β-cap region creates a constraint for the folding of the cluster-binding domain
[16]. As part of the folding process, mitoNEET binds two [2Fe-2S]
2+/+ clusters, one in each subunit of the dimer, using three cysteines (Cys-72, Cys-74, Cys-83) and one histidine (His-87) as ligands (
Figure 1c). The His-87 ligand is located at the N-terminus of the α-helix within the cluster-binding domain and is solvent-accessible, as it is the Fe ion that it coordinates together with Cys-83. The other two ligands (Cys-72 and Cys-74) are, by contrast, buried inside the structure and bind a non-solvent-accessible Fe ion
[14][15][17][14,15,17]. This coordination sphere, that is common to all the NEET proteins, is different from those of ferredoxin-like and Rieske proteins that coordinate [2Fe-2S]
2+/+ clusters either with four cysteines, or with two cysteines and two histidines, respectively. The binding of the [2Fe-2S] clusters to each subunit in the dimer is responsible for the unique fold adopted by the holo protein
[18], and the dimeric arrangement of the CDGSH domains seems to be essential for the stabilization of the coordination sphere of mitoNEET. Indeed it has been proposed that the β-cap domain formed by the strand swapping of the two protomers could function as an allosteric control site, modulating cluster insertion, assembly, or electron transfer
[16]. Even CISD3, considered as the ancestor protein of the family, being monomeric, contains two CDGSH domains in its primary sequence and, as a consequence, two [2Fe-2S] clusters
[19]. No cooperativity effect between the two clusters has been reported so far, even though inter-cluster dipolar coupling has been detected by EPR studies
[20].
MitoNEET takes part into a variety of cellular processes, acting as a regulator of the homeostasis of iron and of reactive oxygen species (ROS)
[21][22][21,22], as well as of the metabolism of glucose and lipids in cells, therefore modulating mitochondrial bioenergetics
[21][23][24][21,23,24]. These functions have been linked to the observed overexpression of mitoNEET in human epithelial breast cancer cells, where the protein has been found to induce tumor cell proliferation, likely maintaining the mitochondrial functions by preventing the accumulation of iron and ROS in the mitochondrial matrix, regulating autophagy signaling, preventing autophagy
[25][26][27][25,26,27]. Recent studies reported that mitoNEET indeed controls the formation and the integrity of inter-mitochondrial junctions and mitochondrial network morphology
[24][28][24,28]. Moreover, it was observed that mitoNEET interacts with the voltage-dependent anion channel 1 (VDAC), thus regulating the free iron level within mitochondria
[29]. This interaction was also proposed to be functional for the maturation of mitoNEET clusters, mediating the interaction between mitoNEET and CISD3 in a process where CISD3 transfers its [2Fe-2S] clusters from inside the mitochondria to mitoNEET
[30]. In addition, mitoNEET was found to be involved in several human pathologies, such as obesity, where the overexpression of mitoNEET enhances lipid accumulation in adipocytes, but preserves insulin sensitivity
[21][31][32][21,31,32] and neurodegeneration
[33]. MitoNEET have been also linked to type 2 diabetes, since it was identified as the main cellular target of the thiazolidinedione (TZD) pioglitazone, a drug extensively used to treat insulin resistance
[7], although the role of mitoNEET in the etiology of the pathology is unclear.
Despite the number of cellular and pathological mitoNEET-related processes, very few details are known about the mechanism of action of the protein in such processes. MitoNEET is supposed to play a major role in repairing the damaged [4Fe-4S] cluster on cytosolic apo aconitase IRP1 in oxidative stress conditions
[18], and acts as a cluster transfer protein for several apo recipient proteins
[18][34][35][18,34,35]. Both functions are based on a redox switch, activated by several cellular components
[36][37][38][39][40][36,37,38,39,40]. Indeed, only [2Fe-2S]
2+ and not [2Fe-2S]
+ clusters can be transferred from holo mitoNEET to apo recipient proteins
[18][34][35][18,34,35]. For this reason, the oxidized and the reduced state of mitoNEET clusters have been defined as “active” and “dormant” states, respectively
[41]. The two [2Fe-2S] clusters of mitoNEET have a ∼0 mV midpoint redox potential in vitro at pH 7.5
[42][43][42,43] and in the cytoplasmic cellular environment, they are in the reduced state, as shown by EPR spectroscopy performed on
E. coli cells containing the overexpressed cytosolic domain of human mitoNEET
[44]. These findings suggest that in normal cellular conditions, mitoNEET clusters are stably bound to the protein in the reduced, dormant state. Several factors can change the redox state, and, therefore, the reactivity of mitoNEET clusters: [2Fe-2S]
2+-mitoNEET can be reduced in vitro by biological thiols
[44], reduced flavin nucleotides
[36][37][38][36,37,38], and enzymes, such as human glutathione reductase
[39]. Additionally, other Fe-S proteins, such as human anamorsin, transfer electrons to mitoNEET in vitro, showing a possible direct link between the cytosolic iron-sulfur cluster assembly (CIA) machinery and the mitoNEET cluster transfer repairing pathway
[40].
Another important factor for the stability/reactivity of mitoNEET is the peculiar pH lability of its [2Fe-2S] clusters. The presence of a His residue in the first coordination sphere of mitoNEET [2Fe-2S] clusters results in a significant sensitivity to pH variations and in the pH lability of the cluster, described as a peculiar feature of the NEET proteins. Indeed, it has been proposed that the protonation of the imidazolic ring of the His-87 ligand at acidic pH facilitates the transfer of the mitoNEET [2Fe-2S]
2+ clusters to apo recipient proteins in vitro, or their release in solution
[18][34][35][41][18,34,35,41]. However, the protonation of the His-87 ligand is likely not the sole factor affecting cluster stability in mitoNEET. Indeed, Rieske proteins, which contain two His residues in their cluster coordination sphere, show a significant higher cluster stability over a wide range of pH
[45]. The pH-dependent stability of mitoNEET clusters seems to be related also to a hydrogen-bonding network formed by the His-87 ligand, a conserved solvent water molecule, and the Nε of Lys-55 residue from the other polypeptide chain of the dimer
[42][46][47][42,46,47]. Moreover, NMR and UV-visible spectroscopies showed that the stability of mitoNEET [2Fe-2S] clusters can be tuned also by the interaction with small molecules, such as the antidiabetic drug pioglitazone, which interacts with the cluster-binding region of the protein, increasing the stability of the two Fe-S clusters
[15][48][15,48], or reduced nicotinamide adenine dinucleotide phosphate (NADPH), which, by contrast, destabilizes mitoNEET Fe-S clusters and induces protein unfolding
[49].
For all the aforementioned characteristics, the study of the electronic and coordination structures of mitoNEET [2Fe-2S] clusters has been attracting increasing interest in the last few years, and, in addition to the above-described X-ray crystallographic studies, a variety of spectroscopic techniques have been applied to study mitoNEET in the attempt to explain the drastic difference in clusters stability and reactivity observed for the two redox states and to clarify the cellular function of the protein. The similarities and the differences between the oxidized and reduced states of the mitoNEET [2Fe-2S] clusters and those of other Fe-S proteins are crucial aspects for understanding their modes of action and their role in the physiological processes.