2. In Vivo Development of Schwann Cells
2.1. Development of Neural Crest and Schwann Cell Precursors
The Schwann cell lineage arises from the neural crest, a multipotent stem cell population specific to vertebrate embryonal development (
Figure 1A). Of ectodermal origin, the neural crest is induced during gastrulation
[24][25][24,25] and subsequently specified from the neural plate border territory by signaling from adjacent developing structures, i.e., the juxtaposed non-neural ectoderm, neural plate, and underlying mesoderm. After closure of the neural tube, premigratory neural crest cells undergo an epithelial-to-mesenchymal transition (EMT), allowing them to delaminate from the neuroepithelium and to migrate through the embryonal tissue. Dorsoventrally emigrating neural crest cells associate with developing peripheral nerves and are then termed Schwann cell precursors
[26][27][26,27]. Both neural crest cells and Schwann cell precursors reach their target tissues through specific migration pathways and differentiate towards their terminal fate once settled in the target tissues, where they often undergo a mesenchymal-to-epithelial transition
[28]. During this migration phase, Schwann cell precursors move along the peripheral nerves.
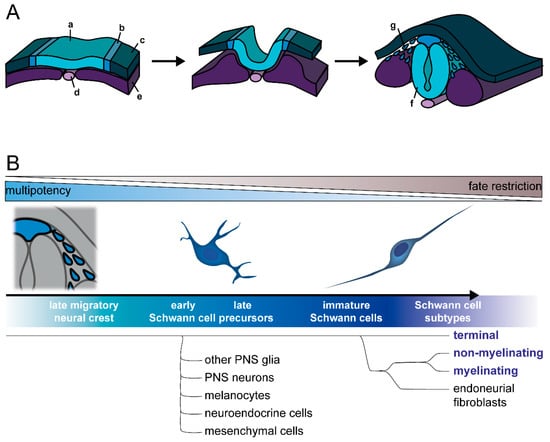
Figure 1. Development of the Schwann cell lineage. (
A) During neurulation, the neural plate border (b) develops between the neural plate (a) and the non-neural ectoderm (c), along neighboring structures, including the notochord (d) and paraxial mesoderm (e). After the neural plate folds to build the neural tube (f), the neural plate borders merge and form the neural crest. Late neural crest cells (g) detach and migrate. (
B) Differentiation along the Schwann cell lineage progresses through several intermediate stages, namely Schwann cell precursors and immature Schwann cells. While Schwann cell precursors are multipotent and can still differentiate into other cell types, such as melanocytes or neurons, immature Schwann cells are more fate restricted and differentiate into mature myelinating and non-myelinating Schwann cells, terminal Schwann cells of the neuromuscular junctions, as well as endoneurial fibroblasts. According to current knowledge, terminal Schwann cells segregate during differentiation prior to endoneurial fibroblasts and other Schwann cell subtypes. Figure from
Hörner et al. 2022 under CC BY 4.0 license.
Neural crest induction and specification are primarily mediated by four major developmental pathways, i.e., Wnt, FGF, BMP, and Notch signaling (for review, see Prasad et al., 2019
[29]). Wnt signaling is crucial for important aspects of neural crest induction, i.e., formation of the neural plate border and subsequent activation of neural crest-specifying transcription factors (reviewed by Ji et al., 2019
[30]). FGF signaling has been especially implicated in the early phases of neural crest specification
[24][31][24,31]. BMP signaling appears to be critical after initial neural crest induction
[32]. It is required at an intermediate activity level, which is provided by gradients of laterally secreted BMP activating signals and medially secreted BMP antagonizing signals
[33][34][35][33,34,35].
Finally, Notch signaling is relevant in neural crest induction, migration, differentiation, and inhibition of neurogenesis
[36][37][38][39][36,37,38,39], but its exact role in these processes and the apparent variations between species remain to be elucidated
[29]. Although a wealth of in vivo and in vitro studies in different species have established these pathways and their crosstalk as principal regulators of neural crest development, new insights about their precise spatiotemporal activities are still emerging. In addition, other pathways have been suggested to play a role in neural crest development, such as retinoic acid signaling
[40][41][40,41] and the Hippo/YAP pathway. For example, recent findings suggest that Hippo/YAP promotes neural crest fate and migration, as well as fate specification of neural crest cells (reviewed by Zhao et al., 2021
[42]): YAP activation could benefit the EMT and migration of neural crest cells, and studies have suggested the crosstalk of YAP signaling with that of BMP and Wnt
[43], retinoic acid
[44], and Notch
[45] during neural crest development.
Overall, an astounding variety of cell types of neural, as well as mesenchymal, fates are generated from the neural crest
[46]. Besides Schwann cells, these also include other peripheral glia such as enteric glia and ganglionic satellite glia, as well as neurons of the peripheral nervous system (PNS), including sensory neurons, postganglionic sympathetic and parasympathetic neurons, and enteric neurons. Furthermore, melanocytes, endocrine cells, and a variety of mesenchymal cells, including cranial bone and connective tissue, are also neural crest descendants. Some of these fates depend on the neural crest anteroposterior axial identity (cranial, vagal, trunk, sacral), as they contribute to different tissue types, suggesting there is already heterogeneity of differentiation competence in the neural crest population
[47][48][47,48]. While neuron and glia cells differentiate from the neural crest at all axial levels, the major part of Schwann cells and PNS neurons are derived from trunk neural crest
[27][49][50][51][52][53][27,49,50,51,52,53].
The development of neural crest cells into mature Schwann cells proceeds through intermediate stages: first, the previously mentioned Schwann cell precursors, which are found in the early embryonic nerve; and then the immature Schwann cells in late embryonic and perinatal nerves (Schwann cell precursor development has been reviewed in detail elsewhere
[1][49][54][55][1,49,54,55]) (
Figure 1B). The extrinsic signals inducing and specifying these stages are still unknown
[54][56][54,56], but a key factor is neuregulin-1 (Nrg1), specifically the axon-derived splicing isoform Nrg1 type III
[57][58][59][57,58,59]. In the Schwann cell precursor stage, membrane-bound Nrg1 type III on juxtaposed sensory and motor axons acts as a crucial survival factor through ErbB family receptor tyrosine kinases, mainly ErbB2/3 heterodimers
[60][61][60,61]. The transition from neural crest to Schwann cell precursors has been connected to the acquisition of typical glia features. In particular, they enter developing nerves, associate with axons, and depend on axonal signals for survival. At the same time, glial and Schwann cell lineage specific markers are upregulated
[1]. Nevertheless, Schwann cell precursors retain a high degree of multipotency, and become primed towards immature Schwann cells or other fates in a gradual manner
[55][62][55,62].
Notably, several neural crest-derived cell types outside the Schwann cell lineage are derived from both the neural crest and Schwann cell precursors. One example is the melanocytes, which differentiate either directly from neural crest cells or from Schwann cell precursors, following their migration along nerves
[63][64][63,64]. A similar mechanism might possibly be true for enteric glia, as both neural crest cells and Schwann cell precursors were found to populate the gut to differentiate
[65]. To date, further cell types which can develop from Schwann cell precursors include endoneurial fibroblasts
[66]; postganglionic parasympathetic neurons
[67][68][67,68]; enteric neurons
[69]; neuroendocrine chromaffin cells of the adrenal medulla
[70] and Zuckerkandl organ
[71], a small population of sympathetic neurons in paraganglia
[71]; dental pulp mesenchymal stromal cells
[72]; and even chondrocytes and osteocytes
[73]; as well as, possibly, recently discovered immune-regulating splenic glia
[74][75][76][74,75,76]. This remarkable developmental potential of multipotent Schwann cell precursors clearly indicates that no terminal commitment to the Schwann cell lineage has happened at this stage. Indeed, recent work using single-cell transcriptomics has confirmed that early Schwann cell precursors are highly similar to late migratory neural crest cells and exhibit multipotency towards various cell fates, while late Schwann cell precursors that progress towards the Schwann cell fate become gradually more restricted
[62].
2.2. Commitment to Schwann Cell Lineage and Differentiation of Subtypes
Commitment to immature Schwann cells follows the multipotent Schwann cell precursor stage (
Figure 1B). After migratory Schwann cell precursors have reached their target tissue, they either detach from nerves and assume other cell fates or further develop into post-migratory immature Schwann cells in a continued association with axons
[55][77][78][55,77,78]. This is accompanied by upregulation of glial and Schwann cell markers (e.g., Krox20, P0, S100b, Oct6) and downregulation of earlier markers (e.g., Ap2α, Pax3, Sox2)
[3][26][79][80][81][3,26,79,80,81]. At the same time, these cells start expressing basal lamina proteins, as well as growth factors, which act on differentiating neurons but also as autocrine survival factors
[1][52][82][83][1,52,82,83]. During this period, immature Schwann cells initiate radial sorting of axons, whereby groups of immature Schwann cells bundle several axons together, before segregating the larger axons designated for myelination; these processes are orchestrated by axonal Nrg1
[84]. Furthermore, Notch signaling seems to be involved in the transition to immature Schwann cells, through boosting Nrg1 signaling; indeed, activation of Notch1 receptor through Jagged1 expressed on adjacent neurons maintains high expression levels of ErbB2 receptors in differentiating Schwann cells. At later stages, Notch inhibits myelination and is suppressed through Krox20 at the onset of the myelination program
[85]. While immature Schwann cells are initially highly proliferative, more than Schwann cell precursors
[86], they gradually exit the cell cycle during maturation, when Schwann cells assume mature myelinating and non-myelinating phenotypes. This mainly occurs postnatally in rodents
[3][86][87][3,86,87].
The two known major subtypes of mature Schwann cells are myelinating Schwann cells and non-myelinating Remak Schwann cells. While the first associate with one individual axon through radial sorting and build up the myelin sheath wrapping a segment of this axon, the latter associate with several small diameter axons, and ensheath them in so-called Remak bundles, without producing myelin. Terminal (or perisynaptic) Schwann cells are a third distinct subtype of Schwann cells. They are associated with neuromuscular junctions, and although they are non-myelinating, terminal Schwann cells differ considerably from Remak Schwann cells in terms of their marker expression, appearance, and function
[7][88][89][7,88,89]. Besides these, it is likely that other glial subtypes that have not been well characterized so far also arise from Schwann cell precursors and might differentiate through the immature Schwann cell stage as well
[79], such as the glia cells of cutaneous sensory corpuscles
[90][91][90,91]. While some factors that promote the development of the mature myelinating Schwann cell subtype are known, the differential mechanisms that specify the mature non-myelinating fates, including Remak and terminal Schwann cells, are largely unclear.
Myelinating Schwann cells develop through a further intermediate state, termed a promyelinating Schwann cell, which establishes a 1:1 relationship with an axon sorted for myelination
[84]. Axonal sorting and maturation of myelinating Schwann cells is dependent on axonal contact and Nrg1 expression levels
[92]. Myelination and terminal differentiation appear to involve activation of the adhesion G-protein coupled receptor GPR126 by basal lamina components such as laminin-211
[93][94][95][96][93,94,95,96]. GPR126 might signal through multiple downstream effectors, one being the myelination onset regulator Krox20
[97] through cAMP-dependent PKA activation
[98]. Furthermore, the axonal Nrg1 expression level seems to tune the regulation of myelination fate. Indeed, while higher Nrg1 levels are required for myelination, lower Nrg1 levels may induce Remak differentiation
[92]. At the same time, the Nrg1 expression level is proportional to the axon diameter, so that large caliber axons are sorted for myelination and small caliber axons are ensheathed by Remak bundles
[99]. In addition, GABA-B receptors were found to regulate development towards non-myelinating Schwann cells through negative modulation of adenylate cyclase and downregulation of myelin-related proteins
[100][101][102][100,101,102]. Furthermore, GPR126 is also involved in Remak Schwann cell differentiation
[95]. However, it is unknown how GPR126 functions mechanistically in this context and how its role differs from that in differentiation of myelinating Schwann cells. As for terminal Schwann cells, to date, no differential specifying molecular factors are known, and even the developmental time window when they diverge from the other subtypes remained unknown until recently. Using single-cell transcriptomics, Kastriti and colleagues analyzed developmental transitions and fate choices along the neural crest and Schwann cell lineages. This revealed that immature Schwann cells develop into four terminally differentiated cell types: terminal, non-myelinating (Remak), and myelinating Schwann cells, as well as endoneurial fibroblasts. Of these, the terminal Schwann cell fate is the first to diverge from the others; thus, even before the path of endoneurial fibroblasts splits from myelinating and Remak Schwann cells (
Figure 1B)
[62].
Even after terminal differentiation into mitotically quiescent mature Schwann cell subtypes, the Schwann cell linage retains a remarkably plastic reprogramming potential, and terminal differentiation is reversible. Predominantly as a reaction to nerve injury, mature Schwann cells of all subtypes can re-enter the cell cycle, de-differentiate, and assume what is called the repair phenotype. This is distinct from other mature, as well as developmental, Schwann cell phenotypes
[12][103][12,103]. Repair Schwann cells are crucial for nerve repair and account for the outstanding regeneration ability of the PNS
[11][104][11,104]. Following the resolution of nerve damage, repair Schwann cells re-differentiate into mature myelinating and non-myelinating Schwann cells. During this re-differentiation process, prior mature myelinating Schwann cells can assume a mature non-myelinating phenotype and vice versa
[105][106][105,106]. Notably, they might even re-differentiate into non-glial cells, including melanocytes
[64], dental pulp mesenchymal stem cells
[72], or enteric neurons
[107]. This remarkable plasticity of developing and mature descendants of the Schwann cell lineage is controlled by transcription factor networks and chromatin remodeling
[108][109][110][108,109,110] and makes Schwann cells exceptionally interesting for in vitro modeling studies on nerve injury, disease, and regeneration. However, their extensive developmental multipotency and adult plasticity also add to the challenges in deriving, characterizing, and maintaining Schwann cells in vitro.