The glutathione system has been recognized as one of the most important intracellular antioxidant systems; however, the contribution of this system in pathogenesis has been neglected, especially in human fungal pathogens.
- glutathione
- metabolism
- morphogenesis
1. Introduction
2. Glutathione Systems
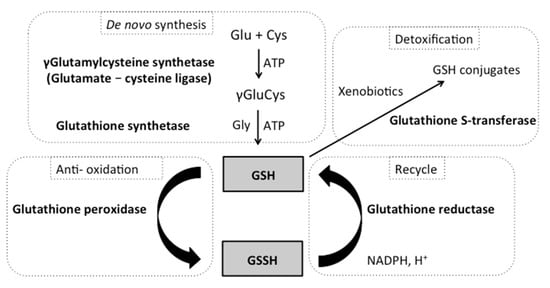
3. Glutathione and Role in Oxidative Stress Protection
Table 1
3.1. γ-Glutamylcysteine Synthetase and Glutathione Synthetase
Table 1. Analyses of glutathione-related enzymes in diverse fungal species.
|
Enzymes in glutathione system |
Species |
Gene name |
Phenotypes |
References |
|
γ-glutamylcysteine synthetase |
Saccharomyces cerevisiae |
GSH1 |
The gsh1Δ mutant showed glutathione auxotrophy, slower growth and increased sensitivity to oxidative stress. |
[28,29] |
|
Schizosaccharomyces pombe |
gcs1 |
- The gcs1Δ mutant showed glutathione auxotrophy and sensitivity to cadmium. - The gcs1Δ mutant was unable to sporulate. |
[30,31] |
|
|
Candida albicans |
GCS1 |
- The gcs1Δ mutant showed glutathione auxotrophy, increased ROS production and apoptosis. - The gcs1Δ mutant showed no change in morphogenesis and virulence. |
[32,33] |
|
|
Nakaseomyces glabrataa (formerly, Candida glabrata) |
GSH1 |
- The gsh1Δ mutant was lethal. - A conditional deletion mutant, gsh1Δpro2-4, showed low glutathione levels and slower growth in media lacking glutathione. - The gsh1Δpro2-4 mutant showed sensitivity to oxidative stress (H2O2, menadione) and cadmium. |
[33,34] |
|
|
Histoplasma capsulatum |
GSH1 |
- The GSH1 gene was expressed only in the yeast form. - The gsh1Δ mutant was lethal. - The GSH1 overexpression mutant showed an inability to switch from yeast to mold form. |
[35] |
|
|
Glutathione synthetase |
Saccharomyces cerevisiae |
GSH2 |
The gsh2Δ and the GSH2 overexpression mutants showed normal responses to oxidative stress. |
[36] |
|
Schizosaccharomyces pombe |
gsh2 |
The gsh2Δ mutant showed glutathione auxotrophy and sensitivity to cadmium. |
[30,31] |
|
|
Nakaseomyces glabrataa (formerly, Candida glabrata) |
GSH2 |
- The gsh2Δ mutant showed glutathione auxotrophy. - The gsh2Δ mutant showed low glutathione levels and sensitivity to oxidative stress (H2O2, menadione) and cadmium. - The gsh2Δ mutant showed resistance to tert-butyl hydroperoxide and cumene hydroperoxide stressors. |
[34] |
|
|
Cryptococcus neoformans |
GSH2 |
- The gsh2Δ mutant showed glutathione auxotrophy under iron starvation conditions. - The gsh2Δ showed low glutathione levels and sensitivity to the oxidative stressor diamide, but not H2O2. - The gsh2Δ mutant showed sensitivity to a high salt stressor, the cell wall damaging agent Congo red, and antifungal drugs. - The gsh2Δ showed impairment in virulence-related traits, including defects in capsule formation, melanin production and growth at 37 °C. |
[24] |
|
|
Histoplasma capsulatum |
GSH2 |
- The GSH2 gene was highly expressed in the yeast form. - The GSH2 overexpression mutant showed an inability to switch from yeast to mold form. - The gsh2Δ mutant was lethal. |
[35] |
|
|
Glutathione reductase |
Saccharomyces cerevisiae |
GLR1 |
The glr1Δ mutant showed sensitivity to oxidative stress (H2O2). |
[37,38] |
|
Schizosaccharomyces pombe |
pgr1 |
- The pgr1 overexpression mutant showed resistance to the oxidative stressor menadione but not H2O2. - The pgr1 gene expression was induced by various oxidative stressors (menadione, cumeme hydroperoxide and diamide, but not H2O2), high salt levels (NaCl), high temperatures and starvation. - The pgr1Δ mutant was lethal. |
[39] |
|
|
Candida albicans |
GLR1 |
- The glr1Δ mutant showed sensitivity to oxidative stress (H2O2) but not formaldehyde or nitrosative stress (NO). - The glr1Δ mutant showed an inability to detoxify GSSG. - The glr1Δ mutant showed impairment in macrophage killing. - The glr1Δ mutant showed decreased virulence, while the GLR1 overexpression mutant showed increased virulence. |
[40] |
|
|
Cryptococcus neoformans |
GLR1 |
- The glr1 gene expression was induced by nitric oxide (NO). - The glr1Δ mutant showed normal morphology. - The glr1Δ mutant showed sensitivity to nitric oxide stress but not peroxide stress. - The glr1Δ mutant became avirulent in an inhalation model of mouse infection and showed sensitivity to macrophage killing. |
[41] |
|
|
Aspergillus nidulans |
glrA |
- The glrAΔ mutant showed slower growth under normal conditions. - The glrAΔ mutant showed defects in conidia germination at high temperatures. - The glrAΔ mutant showed sensitivity to various oxidants (menadione, diamide and H2O2). - The glrAΔ mutant accumulated a less reduced form of GSH, more intracellular ROS, and had decreased respiration activity. |
[42] |
|
|
Paracoccidioides brasiliensis |
GR |
The vPb18 virulent strain showed increases in both levels of the GR gene and enzymatic activity. |
[43] |
|
|
Glutathione peroxidase |
Saccharomyces cerevisiae |
GPX1-3 |
- GPX1-3 genes encoded for phospholipid hydroperoxide glutathione peroxidase. - The GPX3 product was a major glutathione peroxidase. - The GPX3 gene was constitutively expressed. - The GPX1 gene expression was induced under glucose starvation. - The GPX2 gene expression was induced by many oxidative stressors. - The gpx3Δ mutant showed sensitivity to peroxides (H2O2 and tert-butyl hydroperoxide). - The gpx1Δ and gpx2Δ mutants showed no sensitivity to oxidative stress. - The gpx1Δgpx2Δgpx3Δ mutant showed sensitivity to H2O2 and phospholipid hydroperoxide (polysaturated fatty acid linolenate 18:3). |
[44,45] |
|
Candida albicans |
GPX3 (ScGPX1 homolog) |
- The gpx3Δ mutant (orf19.4436Δ) showed sensitivity to H2O2 and was defective in hyphal formation within macrophage cells. - The gpx3Δ mutant showed impairment in killing macrophages and Galleria mellonella. - The gpx3Δ mutant showed normal virulence in a murine model of infection. |
[46,47] |
|
|
GPX31- 33 (ScGPX3/ HYR1 homolog |
- The GPX31 is a major glutathione peroxidase. - The gpx31Δ (orf19.86Δ) and the gpx31Δgpx32Δgpx33Δ mutant (orf19.86Δorf19.85Δorf19.87Δ) showed sensitivity to oxidative stressors (H2O2 and t-butylhydroperoxide but not menadione), UV light, heavy metals (cadmium and silver), and cell wall stressors (congo red and calcofluor white). |
|||
|
Cryptococcus neoformans |
GPX1, GPX2 |
- GPX1 and GPX2 gene expressions were induced in response to t-butylhydroperoxide and cumene hydroperoxide and repressed in response to nitric oxide. - GPX2 gene expression was induced in response to the hydrogen peroxide stressor. - The gpx1Δ and gpx2Δ mutants showed normal morphology, melanin production and capsule formation. - The gpx1Δ and gpx2Δ mutants showed sensitivity to cumene (hydroperoxide) but not superoxide, hydrogen peroxide or nitric oxide stressors. - The gpx2Δ mutant showed higher sensitivity to cumene hydroperoxide than the gpx1Δ mutant at high concentrations. - The gpx1Δ mutant, but not the gpx2Δ mutant, showed sensitivity to the peroxide stressor t-butylhydroperoxide. - The gpx1Δ and gpx2Δ mutants showed sensitivity to macrophage killing, yet the mutants were still virulent in a mouse model. |
[48] |
|
|
Aspergillus fumigatus |
hyr1 (ScGPX3/ HYR1 homolog) |
- hyr1 gene expression was upregulated in hyphae and conidia when exposed to neutrophils. - The hyr1 gene expression was induced when exposed to H2O2. |
[49,50] |
|
|
Talaromyces marneffei |
gpx1 (ScGPX3/HYR1 homolog) |
- Gpx1 is an antigenic protein. - gpx1 gene expression was upregulated in the yeast form. |
[51,52] |
|
|
Glutathione S- transferase |
Saccharomyces cerevisiae |
GTT1-2 |
- GTT1 gene expression was induced during the diauxic shift and stationary phase. - The gtt1Δ, gtt2Δ, and gtt1Δgtt2Δ showed sensitivity to heat shock in a stationary phase and slower growth at a high temperature of 39 °C. - The grx1Δgrx2Δgtt1Δgtt2Δ mutant showed sensitivity to xenobiotics (1-chloro-2,4-dinitrobenzene), heat and the oxidative stressors (cumene hydroperoxide and H2O2). |
[53,54] |
|
Schizosaccharomyces pombe |
gst1-3, |
- The gst1Δgst2Δ and gst3Δ mutants showed sensitivity to peroxide stressors (H2O2 and t-butylhydroperoxide) and the antifungal drug fluconazole. - The gst1Δgst2Δ and gst3Δ mutants showed resistance to the peroxide stressor diamide. - gst1, gst2, and gst3 gene expressions were induced during the stationary phase and in response to hydrogen peroxide. - All Gst1, 2 and 3 enzymes have glutathione transferase activity, and the Gst3 enzyme also has glutathione peroxidase activity. |
[55] |
|
|
Candida albicans |
GST2 |
- The gst2Δ mutant showed sensitivity to oxidative stress (H2O2). - GST2 gene expression was induced under nitrogen limitation. - The gst2Δ mutant showed defects in hyphal switching under nitrogen starvation-induced filamentous growth. |
[56] |
|
|
Aspergillus nidulans |
gstA |
- The gstAΔ mutant showed sensitivity to the oxidant diamide, the fungicide carboxin, various xenobiotics (pyrrolnitrin and sulphanilamide), and heavy metals (selenium, silver and nickel). - The gstAΔ mutant showed normal growth in the presence of 1-chloro-2,4-dinitrobenzene. - The gstA gene expression was induced in response to xenobiotics (1-chloro-2,4-dinitrobenzene) and oxidative stress (H2O2). |
[57] |
|
|
Aspergillus fumigatus |
gstA, gstB, gstC |
- All gstA, B and C enzymes have both glutathione transferase and glutathione peroxidase activities. - The gstA and gstC genes were constitutively expressed under normal conditions, and their expression levels were inducible in response to oxidative stress (H2O2). - The expression of all gst genes was induced in response to xenobiotics (1-chloro-2,4-dinitrobenzene). |
[58] |
|
|
Paracoccidioides brasiliensis |
GST1-3 |
The vPb18 virulent strain showed increased levels of the GST1-3 genes. |
[43] |
|
|
Paracoccidioides lutzii |
GST |
GST was exclusively secreted in the yeast form. |
[59] |
1. Introduction
2. Glutathione Systems
3. Glutathione and Role in Oxidative Stress Protection
3.1. γ-Glutamylcysteine Synthetase and Glutathione Synthetase
3.2. Glutathione Reductase
3.3. Glutathione Peroxidase
3.4. Glutathione S-Transferase
References
- Brown, A. Integration of metabolism with virulence in Candida albicans. In Fungal Genomics; Springer: Berlin/Heidelberg, Germany, 2006; pp. 185–203.
- Brown, A.J.; Brown, G.D.; Netea, M.G.; Gow, N.A. Metabolism impacts upon Candida immunogenicity and pathogenicity at multiple levels. Trends Microbiol. 2014, 22, 614–622.
- Brown, G.D.; Denning, D.W.; Gow, N.A.; Levitz, S.M.; Netea, M.G.; White, T.C. Hidden killers: Human fungal infections. Sci. Transl. Med. 2012, 4, 165rv13.
- Kim, J.Y. Human fungal pathogens: Why should we learn? J. Microbiol. 2016, 54, 145–148.
- Gilbert, A.S.; Wheeler, R.T.; May, R.C. Fungal pathogens: Survival and replication within macrophages. Cold Spring Harb. Perspect. Med. 2014, 5, a019661.
- Gross, N.T.; Nessa, K.; Camner, P.; Jarstrand, C. Production of nitric oxide by rat alveolar macrophages stimulated by Cryptococcus neoformans or Aspergillus fumigatus. Med. Mycol. 1999, 37, 151–157.
- Hidalgo, H.A.; Helmke, R.J.; German, V.F.; Mangos, J.A. Pneumocystis carinii induces an oxidative burst in alveolar macrophages. Infect. Immun. 1992, 60, 1–7.
- Irato, P.; Santovito, G. Enzymatic and Non-Enzymatic Molecules with Antioxidant Function. Antioxidants 2021, 10, 579.
- Edwards, J.A.; Chen, C.; Kemski, M.M.; Hu, J.; Mitchell, T.K.; Rappleye, C.A. Histoplasma yeast and mycelial transcriptomes reveal pathogenic-phase and lineage-specific gene expression profiles. BMC Genom. 2013, 14, 695.
- Nemecek, J.C.; Wüthrich, M.; Klein, B.S. Global control of dimorphism and virulence in fungi. Science 2006, 312, 583–588.
- Beyhan, S.; Gutierrez, M.; Voorhies, M.; Sil, A. A temperature-responsive network links cell shape and virulence traits in a primary fungal pathogen. PLoS Biol. 2013, 11, e1001614.
- Noble, S.M.; Gianetti, B.A.; Witchley, J.N. Candida albicans cell-type switching and functional plasticity in the mammalian host. Nat. Rev. Microbiol. 2017, 15, 96–108.
- Tochigi, N.; Okubo, Y.; Ando, T.; Wakayama, M.; Shinozaki, M.; Gocho, K.; Hata, Y.; Ishiwatari, T.; Nemoto, T.; Shibuya, K. Histopathological implications of Aspergillus infection in lung. Mediat. Inflamm. 2013, 2013, 809798.
- Mah, J.H.; Yu, J.H. Upstream and downstream regulation of asexual development in Aspergillus fumigatus. Eukaryot. Cell 2006, 5, 1585–1595.
- Stewart, J.I.P.; Fava, V.M.; Kerkaert, J.D.; Subramanian, A.S.; Gravelat, F.N.; Lehoux, M.; Howell, P.L.; Cramer, R.A.; Sheppard, D.C. Reducing Aspergillus fumigatus virulence through targeted dysregulation of the conidiation pathway. mBio 2020, 11, e03202-19.
- Boyce, K.J.; Andrianopoulos, A. Fungal dimorphism: The switch from hyphae to yeast is a specialized morphogenetic adaptation allowing colonization of a host. FEMS Microbiol. Rev. 2015, 39, 797–811.
- Lo, H.J.; Köhler, J.R.; DiDomenico, B.; Loebenberg, D.; Cacciapuoti, A.; Fink, G.R. Nonfilamentous C. albicans mutants are avirulent. Cell 1997, 90, 939–949.
- Thompson, D.S.; Carlisle, P.L.; Kadosh, D. Coevolution of morphology and virulence in Candida species. Eukaryot. Cell 2011, 10, 1173–1182.
- Carlos, I.Z.; Silva Monnazzi, L.G.; Falcão, D.P.; Machado de Medeiros, B.M. TNF-alpha, H2O2 and NO response of peritoneal macrophages to Yersinia enterocolitica O:3 derivatives. Microbes Infect. 2004, 6, 207–212.
- Penninckx, M.J. An overview on glutathione in Saccharomyces versus non-conventional yeasts. FEMS Yeast Res. 2002, 2, 295–305.
- Grant, C.M. Role of the glutathione/glutaredoxin and thioredoxin systems in yeast growth and response to stress conditions. Mol. Microbiol. 2001, 39, 533–541.
- Pizzorno, J. Glutathione! Integr. Med. (Encinitas) 2014, 13, 8–12.
- Misslinger, M.; Lechner, B.E.; Bacher, K.; Haas, H. Iron-sensing is governed by mitochondrial, not by cytosolic iron-sulfur cluster biogenesis in Aspergillus fumigatus. Metallomics 2018, 10, 1687–1700.
- Attarian, R. Analysis of the Roles of a Monothiol Glutaredoxin and Glutathione Synthetase in the Virulence of the AIDS-Associated Pathogen Cryptococcus neoformans. Ph.D. Thesis, University of British Columbia, Vancouver, BC, Canada, 2016.
- Kumar, C.; Igbaria, A.; D’Autreaux, B.; Planson, A.G.; Junot, C.; Godat, E.; Bachhawat, A.K.; Delaunay-Moisan, A.; Toledano, M.B. Glutathione revisited: A vital function in iron metabolism and ancillary role in thiol-redox control. EMBO J. 2011, 30, 2044–2056.
- Berndt, C.; Lillig, C.H. Glutathione, glutaredoxins, and iron. Antioxid. Redox Signal. 2017, 27, 1235–1251.
- Hattori, R.; Tada, S.; Matsushita-Morita, M.; Suzuki, S.; Kusumoto, K.-I. Gamma-glutamylcysteine synthetase gene homolog (gshA) isiImportant in glutathione homeostasis in Aspergillus oryzae. Jpn. Agric. Res.Q. JARQ 2018, 52, 301–305.
- Lee, J.; Dawes, I.W.; Roe, J.H. Isolation, expression, and regulation of the pgr1(+) gene encoding glutathione reductase absolutely required for the growth of Schizosaccharomyces pombe. J. Biol. Chem. 1997, 272, 23042–23049.
- Song, J.Y.; Cha, J.; Lee, J.; Roe, J.H. Glutathione reductase and a mitochondrial thioredoxin play overlapping roles in maintaining iron-sulfur enzymes in fission yeast. Eukaryot. Cell 2006, 5, 1857–1865.
- Collinson, L.P.; Dawes, I.W. Isolation, characterization and overexpression of the yeast gene, GLR1, encoding glutathione reductase. Gene 1995, 156, 123–127.
- Grant, C.M.; Collinson, L.P.; Roe, J.H.; Dawes, I.W. Yeast glutathione reductase is required for protection against oxidative stress and is a target gene for yAP-1 transcriptional regulation. Mol. Microbiol. 1996, 21, 171–179.
- Tillmann, A.T.; Strijbis, K.; Cameron, G.; Radmaneshfar, E.; Thiel, M.; Munro, C.A.; MacCallum, D.M.; Distel, B.; Gow, N.A.; Brown, A.J. Contribution of Fdh3 and Glr1 to Glutathione Redox State, Stress adaptation and virulence in Candida albicans. PLoS ONE 2015, 10, e0126940.
- Missall, T.A.; Pusateri, M.E.; Donlin, M.J.; Chambers, K.T.; Corbett, J.A.; Lodge, J.K. Posttranslational, translational, and transcriptional responses to nitric oxide stress in Cryptococcus neoformans: Implications for virulence. Eukaryot. Cell 2006, 5, 518–529.
- Sato, I.; Shimizu, M.; Hoshino, T.; Takaya, N. The glutathione system of Aspergillus nidulans involves a fungus-specific glutathione S-transferase. J. Biol. Chem. 2009, 284, 8042–8053.
- Bakti, F.; Király, A.; Orosz, E.; Miskei, M.; Emri, T.; Leiter, É.; Pócsi, I. Study on the glutathione metabolism of the filamentous fungus Aspergillus nidulans. Acta Microbiol. Immunol. Hung. 2017, 64, 255–272.
- Pongpom, M.; Vanittanakom, N. Stress adaptation in Talaromyces marneffei. Chiang Mai Med. J. 2016, 55, 23–30.
- Lee, S.H.; Chung, S.C.; Shin, J.; Oh, K.B. GST2 is required for nitrogen starvation-induced filamentous growth in Candida albicans. J. Microbiol. Biotechnol. 2014, 24, 1207–1215.
- Fraser, J.A.; Davis, M.A.; Hynes, M.J. A gene from Aspergillus nidulans with similarity to URE2 of Saccharomyces cerevisiae encodes a glutathione S-transferase which contributes to heavy metal and xenobiotic resistance. Appl. Environ. Microbiol. 2002, 68, 2802–2808.