1. Introduction
Melatonin (MEL)
N-acetyl-5-methoxytryptamine is a hormone synthesized by the pineal gland. Discovered and isolated from bovine pineal glands by Aaron Lerner, MEL was injected in Lerner’s groundbreaking experiment into dermatosis patients who, contrary to his expectations, did not experience depigmentation but drowsiness. With these initial findings, he began to investigate MEL
[1]. Later, MEL was described in plants as phytomelatonin
[2][3][4][2,3,4], and, nowadays, it is known that MEL is present in practically all organisms along the phylogenetic scale
[5].
The pineal gland produces MEL in the dark phase of the photoperiod. MEL is secreted into the general circulation and in the cerebrospinal fluid and, in this manner, circulates throughout the body and the brain
[6]. Its synthesis is regulated by a clock located in the suprachiasmatic nuclei (SCN) at the hypothalamus. This clock determines the circadian rhythm of MEL secretion. In the brain, extrapineal MEL behaves similarly to Neurotrophic Factors (NTFs)
[7][8][7,8] and is capable of modulating cell survival, proliferation, and differentiation, by signaling pathways that can be triggered in response to stimulation of membrane and intracellular receptors.
In this regard, NTFs play a crucial role in brain neuroplasticity and neurodevelopment, promoting its growth and survival
[9][10][9,10]. These peptide molecules function as signals that trigger biological processes allowing adaptation to the environment and survival of neurons. In the early stages of development, they modulate both differentiation and maturation of neuronal precursors. Some factors are only present at the early stages of development and others throughout life
[10][11][12][10,11,12]. Besides neurogenesis, NTFs are also needed for the maintenance of neuronal function and the neuron’s structural integrity
[11][13][11,13]. NTFs are also expressed in non-neuronal tissues like lung components including nasal and bronchial epithelium, smooth muscle, nerves, immune cells, kidney, spermatozoa, and ovarium
[9][14][15][9,14,15]. These peptides are secreted into the cellular medium and may act in a paracrine manner signaling neighboring cells. If required, cells can also synthesize and secrete these factors in response to autocrine stimulation
[16].
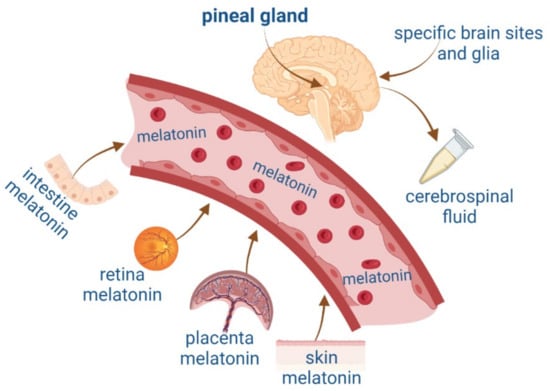
MEL can also be synthesized in other organs such as the intestine, retina, placenta, specific brain regions, and skin
[17][18][17,18] (
Figure 1). MEL is released at micromolar concentrations from extrapineal sites of synthesis and has paracrine, autocrine, and antioxidant actions
[19]. The pleiotropic effects of indolamine have been widely described.
Figure 1.
Melatonin synthesis in the pineal gland and extrapineal sites such as the intestine, retina, placenta, and skin and their blood circulation. Figure created by
.
2. Melatonin and Neurotrophic Factors Synthesis
MEL is an indolamine with a 3-amido group and a 5-methoxy group, which confers its amphiphilic properties to MEL (
N-[2-(5-methoxy-1
H-indol-3-yl)ethyl] ethanamide)
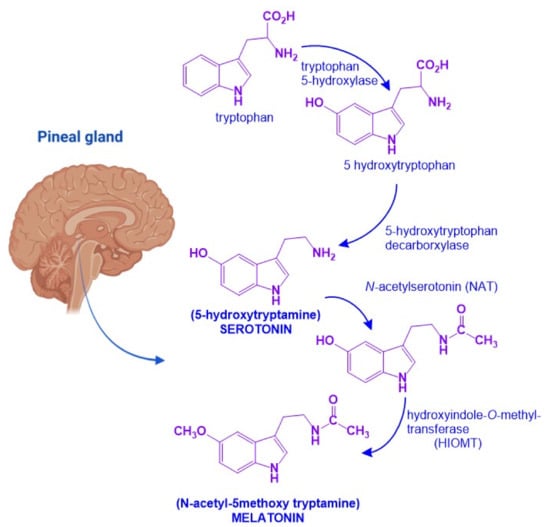
[20]. MEL biosynthesis starts from the amino acid tryptophan which is transformed into serotonin. In the pineal gland, serotonin will undergo two enzymatic reactions: First,
N-acetylation by
N-acetytransferase (NAT) will produce
N-acetyl-serotonin. Then, a methyl group is transferred from
S-adenosylmethionine to the 5-hydroxy group of
N-acetylserotonin by the action of hydroxyindol-
O-methyltransferase (HIOMT)
[21]. NAT is the limiting step for MEL synthesis, and it has a high amplitude rhythm with a nocturnal activity that is 50 to 100 times greater than diurnal activity. HIOMT, on the other hand, catalyzes the last step of the synthesis and has a very low rhythmic amplitude (
Figure 2)
[21].
Figure 2. Melatonin biosynthetic pathway. Melatonin biosynthesis in the pineal gland starts from tryptophan and involves four sequential enzymatic steps to render 5-hydroxytryptophan,
N-acetylserotonin, 5-hydroxytryptamine (serotonin), and
N-acetyltryptamine (melatonin). Figure created with
Biorender.com software.
MEL in circulation is primarily catabolized in the liver and the brain. In the hepatocytes, MEL undergoes 80% of hydroxylation by the cytochrome p450 1A2 (CYP1A2); afterward, in the kidney, it is sulpho-conjugated (70 to 80% of total catabolism) or conjugated with glucuronic acid (5% of total catabolism)
[22]. CYP1A2 ensures hepatic catabolism
[23]. Even at the hepatic level, there are also deacetylation and demethylation pathways. After deacetylation, MEL is transformed into 5-methoxytryptamine and after oxidation to 5-methoxyindole acetic acid
[24]. Cleavage of the pyrrole nucleus of MEL results in
N-acetyl-5-methoxykynurenamine through the
N-formyl intermediate
[24]. 70% of this metabolite is eliminated by urine and 20% in the feces in 24 h. The main route of elimination is, therefore, urinary and will lead to the formation of various metabolites present in the urine: 6-hydroxymelatonin in the form of sulfate (70–80%) and glucuronide (5%) and kynuramines (15%), among them. In summary, MEL urinary elimination comprises MEL (1%), 5-methoxyindoleacetic acid (0.5%), and 6-sulfatoxymelatonin, which is the major urinary metabolite
[25].
On the other hand, NTFs have significant differences in their biosynthesis and catabolism. MEL is an indolamine, while NTFs are peptides synthesized from protein precursors. Despite the great number of molecules considered as NTFs that can regulate neuronal function, the researchers will focus on just four of them with similar structure and functions: Nerve Growth Factor (NGF), Brain-Derived Neurotrophic Factor (BDNF), Neurotrophin-3 (NT3), and Neurotrophin-4/5 (NT4/5).
NTFs are synthesized by neuronal cells as inactive precursors (pro-NTFs, 27 kDa). Synthesis of Pro-NTFs occurs in the endoplasmic reticulum and then a cleavage is caused by furin enzymes in the intracellular compartment to produce active molecules. Also, cleavage is produced by plasmin metalloproteinases in the extracellular space. Pro-NTFs are further processed to generate mature NTFs nearly 13–15 kDa polypeptides with extensive homology. The mature forms of BDNF, NT4, and NT3 have approximately 50% amino acid identity to NGF
[9][26][9,26]. Mature NTFs form homodimers by noncovalent binding and contain a conserved terminal fold and a cysteine “knot”, which consists of three disulfide bonds in the polypeptide chain
[16][27][16,27].
3. The Neurodevelopment in the Adult Brain
The organisms could survive and adapt to their environment through neuroplastic changes produced in the brain. Neuronal activity can modify the number and strength of synaptic connections
[28]. The activity-dependent modulation of synapses is a critical factor for brain development alongside many cognitive functions and behavior. It has been postulated that from the embryonic stages and during adulthood, various cytotypes, tissues, and organs release NTFs. These factors are essential for new neuron formation, their survival as well as differentiation, and the correct organization of the nervous system
[29].
NTFs expression is regulated by neuronal activity and these molecules modulate the efficiency of synaptic transmission, dendrite, and axon growth, as well as the elements necessary for synaptogenesis
[30]. These processes are adaptative changes in which the number of neurons correlates with needs and the number of neurons in the innervated target neurons. The “
neurotrophic factor hypothesis” was conceptualized by Victor Hamburger and Rita Levi Montalcini, who identified the first of these factors in NGF
[31]. In the adult mammalian brain, there are several neurogenic niches: the hippocampal dentate gyrus, the subventricular zone of the lateral cerebral ventricle, the substantia nigra and the cerebellum, the amygdala, the spinal cord, the hypothalamus, and the cerebral neocortex
[32][33][32,33].
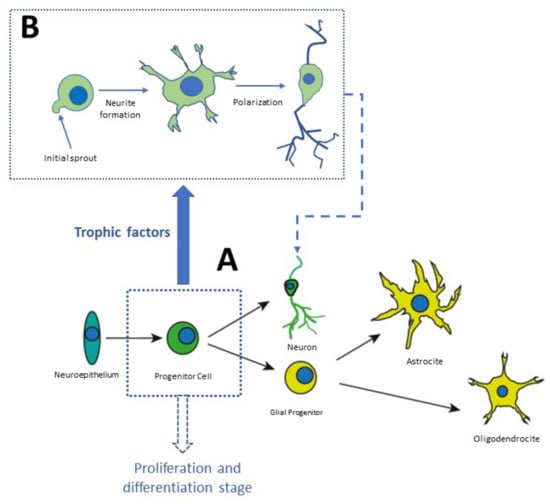
Neurogenesis and neuronal differentiation take place at three stages: (1) Nervous cell generation through cellular proliferation, (2) Migration of neuronal precursor to the area where they will be established, (3) Morphofunctional polarization of neurons in the somatodendritic and axonal domains
[34]. Communication between distal neurons occurs in the third stage of neurogenesis, in which axons and dendrites are formed, synapses are formed according to electric activity patterns, and nonfunctional connections are eliminated
[34]. This process begins with the neuronal secretion of neurotrophic factors. The nerve terminals of neighbor neurons capture these factors, internalize them, and transport them in a retrograde direction to the soma. Growth factors are constantly secreted to establish functional connections, as shown in
Figure 3. Finally, the neurons that are not exposed to the secreted NTFs enter apoptosis and die (
Figure 3).
Figure 3. Stages of neurodevelopment in vitro. Stem cells derived from the neuronal niches located at specific regions in the brain or in the olfactory neuroepithelium can be plated in Petri dishes. In culture, they proliferate and amplify the population of progenitor cells which differentiate into neurons or glial cells, as shown in panel (
A). Panel (
B) shows distinct phases of differentiation
[35][36][35,36].
4. Evidence That Supports Melatonin Acts as a Neurotrophic Factor
Like the neurotrophic factors, MEL also participates in brain neuroplasticity and neurodevelopment
[37][38][37,38]. One important feature of neurogenesis is the proliferation and survival of neuronal precursors. MEL in vitro at 10
−6 M stimulates both processes
[39]. Moreover, with MEL at 10
−7 M, the formation of new neurons increased by approximately 70% in comparison to the control. On the other hand, the indolamine administered for 14 days to mice increases the number of “newborn” neurons in the dentate gyrus of the hippocampus
[40]. However, there was no increase in the proliferation rate. Instead, an increase in the survival of neuronal precursors was observed during the proliferation process. By contrast, other authors observed increased proliferation of new neurons (158%) in the subventricular zone of the adult mouse brain and rat embryos, with concentrations ranging between 10
−7 M and 10
−6 M
[39][41][39,41]. These effects were dose-dependent because higher MEL concentrations increased cell survival
[39][41][39,41]. Environmental factors in combination with MEL also promote neuroplastic changes, for instance, physical activity plus MEL administration enhances neurogenesis in mice
[42].
Recently, the researchers found increased neurogenesis in human olfactory neuronal precursors incubated with MEL, ketamine, and a combination of these compounds. This effect was similar to what has been observed in relation to neurotrophic factors BDNF, Epithelial Grow Factor (EGF), and Fibroblast Growth Factor (FGF) in the dentate gyrus of the hippocampus. The increased neurogenesis was associated with the antidepressant-like behavior produced by these NTFs and the combination MEL/KET
[43][44][43,44].
MEL also stimulates dendritogenesis, dendrite’s spine formation, dendritic arborization, and synaptogenesis
[37][45][37,45]. The indolamine maintains the neuronal somatodendritic domain
[46] through structural morphofunctional polarization thanks to cytoskeletal rearrangements. The three cytoskeletal components (microtubules, microfilaments, and intermediate filaments) are reorganized in the presence of this indolamine
[46]. Hence, an important function of MEL for neurodevelopment is the modulation of the cytoskeletal organization. Cytoskeletal rearrangements play a crucial role in the formation and enlargement of axons and dendrites, as well as in the synaptic assembly (for review see
[46][47][46,47]).
Regarding dendritogenesis, there is a vast literature about the effects of MEL in dendrite formation in organotypic cultures of hippocampus and animal models. Besides the survival-promoting effect, MEL also increases new neuron maturation in adult brains
[48], by augmenting the ramifications of the dendritic trees in the hilus of the hippocampus. In this sense, the systemic administration of MEL to mice for 14 days produced an increase in the number of dendritic arborizations. Of utmost importance is that the loss of dendrites in epilepsy and Alzheimer’s (AD) diseases also occurs in this brain region
[45][49][45,49] and that enhanced dendritic complexity, which is measured in arborizations in the dorsal-ventral regions of the dentate gyrus in male Balb/C mice, is associated with an antidepressant-like behavior
[50]. MEL has also a neuroprotective effect on dendrite formation, for example, administration of MEL to rats, which were previously submitted to global ischemia, prevents the impairment of place learning and memory, both of which are integrated into the hippocampus. Just as importantly, MEL partly preserves the density of spines, mushroom spines, and dendrites pyramidal neurons, which are necessary for an adequate synapsis formation
[51]. The indolamine not only counteracted and protected the neurons against apoptosis; it also prevented and reversed the dendritic arborization retraction
[52], and fostered synapse formation in hippocampal organotypic cultures at 100 nM. This was evidenced by staining with an anti-synapsin antibody which labels synapsin, a protein localized in the presynapsis
[45].
The evidence that supports the stimulation of axogenesis by MEL was obtained in cultured neurons and brain tissue. In cultured human olfactory neuronal precursors, MEL at 10
−7 and 10
−5 M increased by 15% the axonal formation
[37]. Moreover, neurite formation—the primary neurodevelopment step that antecedes axogenesis—is stimulated by MEL in N1E-115 cells
[47]. In addition, experiments in rodents support that MEL stimulates axogenesis.
4.1. Mechanism of Action Involved in Neurogenesis and Neural Differentiation: Neurotrophic Factors and Melatonin
Mechanism of Action Involved in Neurogenesis and Neural Differentiation: Neurotrophic Factors and Melatonin
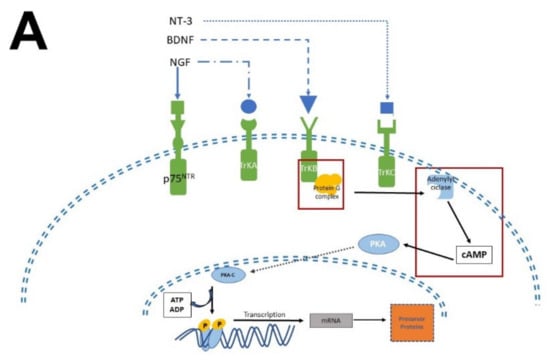
The biological activity of NTFs and MEL occurs thanks to their interaction with transmembrane receptor proteins. In the case of NTFs receptors, they have an extracellular domain, where the NTFs bind
[12][26][12,26], and a cytosolic domain with catalytic and regulatory activity. In contrast, MEL receptors have seven transmembrane domains with an extracellular N-terminal and an intracellular C-terminal
[53][54]. Given that the transactivation of Trk receptors is well documented
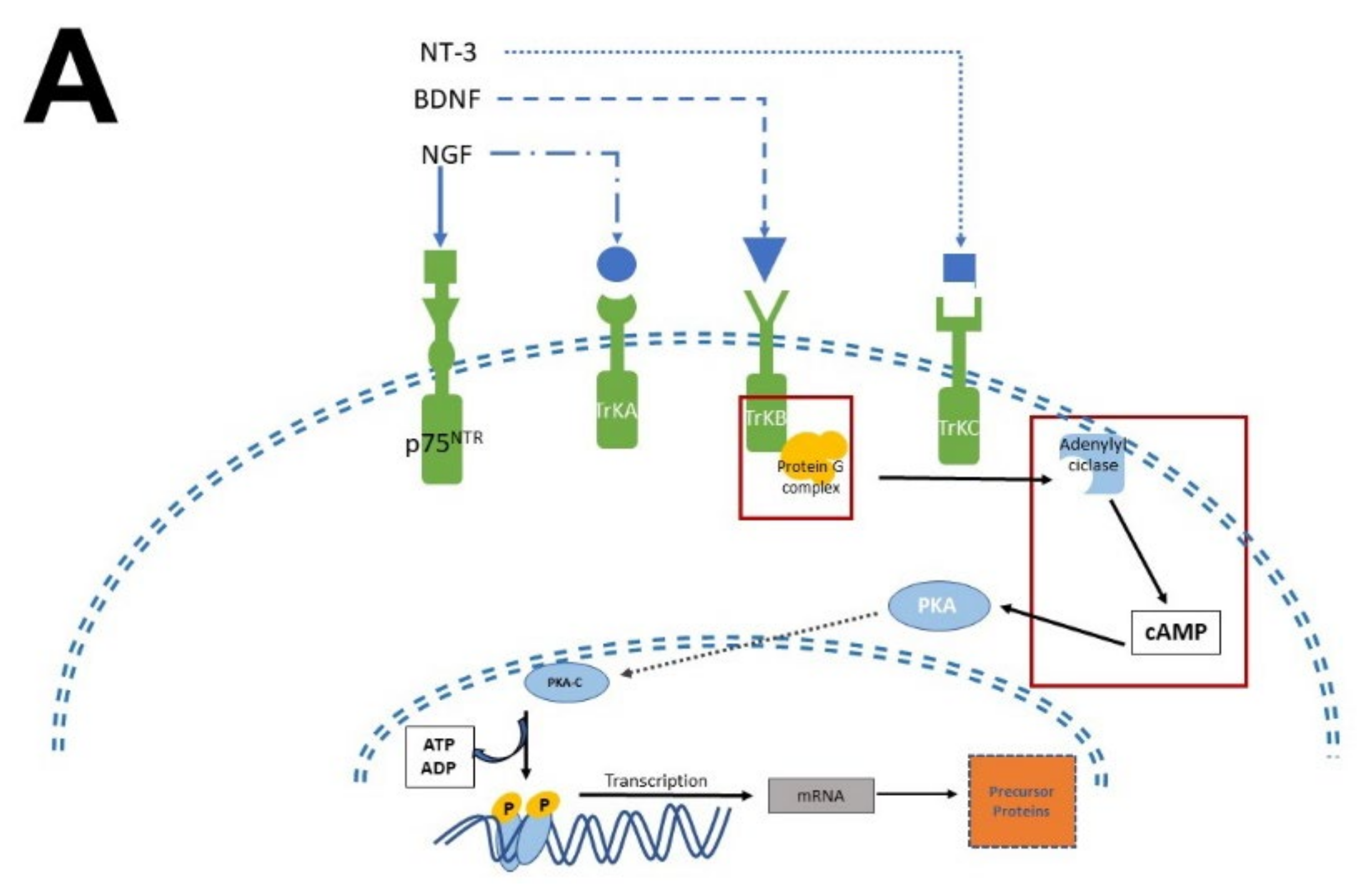
[54][55][55,56], the pathways involved in neurogenesis, proliferation, and survival will be briefly described. MEL participates in all these stages (
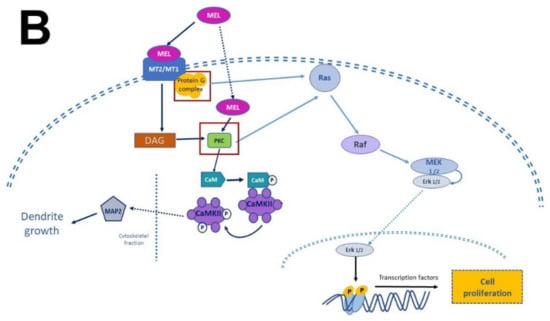
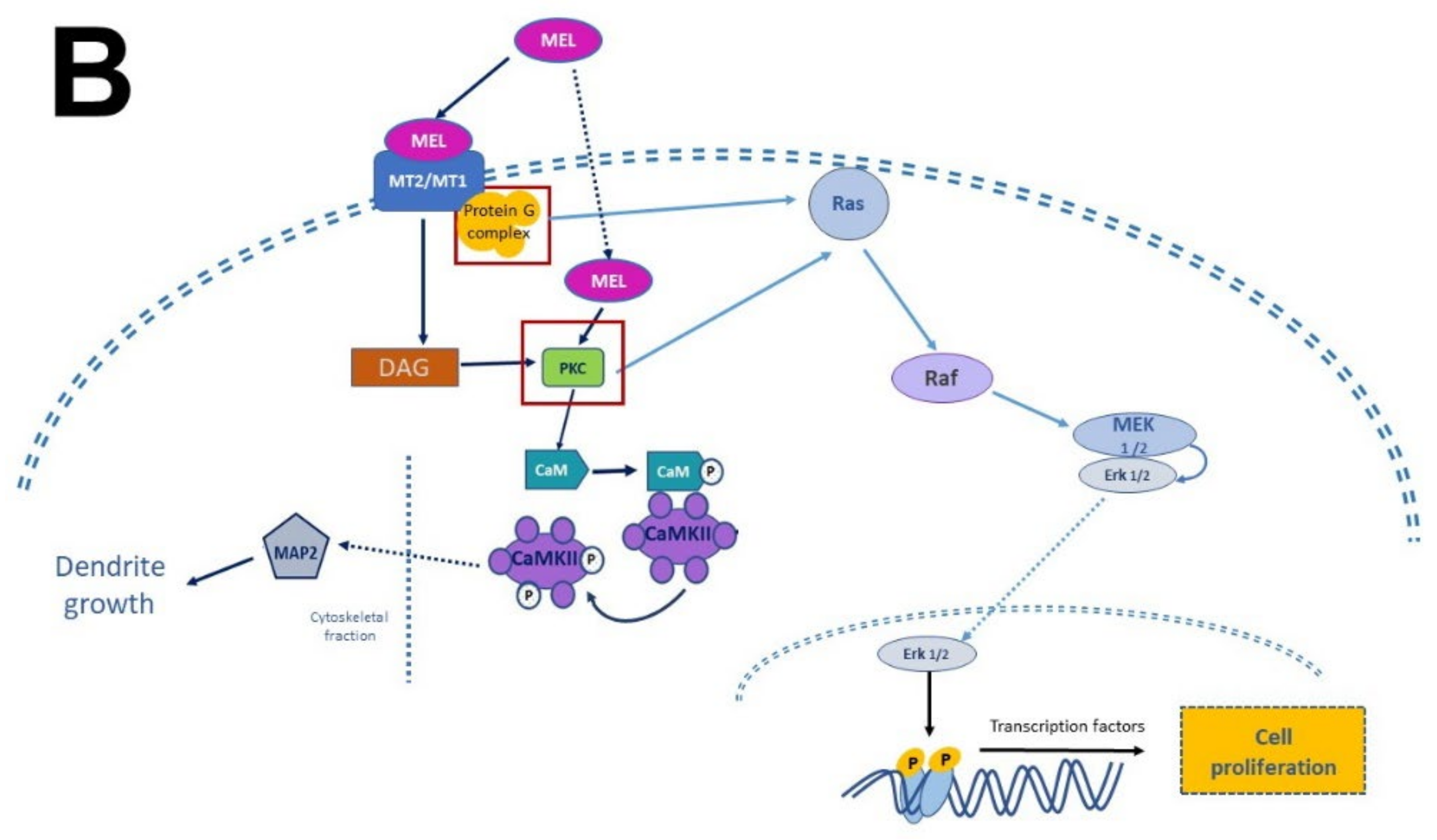
Figure 4).
Figure 4. Simplified scheme of neurotrophic factors and melatonin signaling pathways. Panel (A) shows the interaction of neurotrophins (NT-3, BDNF, NGF) with their receptors (p75NTR, TrkA, TrkB, TrkC), the activation of G proteins, which in turn activate protein kinase A (PKA) and protein kinase C (PKC) to generate transcription factors and precursor proteins that promote neuronal survival. Panel (B) display melatonin (MEL) signaling pathways that promote cell proliferation through binding to melatonin receptors (MT1, MT2), which activate the protein G complex and fibrosarcoma kinases (Raf), mitogen-activated protein kinase (MEK) and extracellular signal-regulated kinase (Erk). MEL can also cross plasmatic membranes and bind to calmodulin (CaM); the complex CaM/MEL binds to the enzyme calmodulin kinase II and enhance its activity to stimulate dendritic development and growth. Red squares in both panels indicate the common features in these pathways.
There are two types of receptors for NTFs: a low NTFs affinity pan-NTF receptor p75NTR that belongs to Tumoral Necrosis Factor (TNF) family, and the high-affinity tropomyosin-related kinase (Trk) receptors; both activate multiple signaling pathways
[26][29][55][56][26,29,56,57]. NGF preferentially activates TrkA, BDNF, and NT-4 activates TrkB, while NT-3 activates TrkC
[57][58]. Once NTFs are bound, the receptor dimerizes and phosphorylates itself on the cytoplasmic domain. When the receptor is phosphorylated, it forms the core of the binding site, adapting proteins and enzymes that mediate the rapid (seconds to minutes) activation of downstream signaling cascades. The main signaling pathways activated by Trk receptors are Ras, phosphoinositide-3 kinase (PI3K), and phospholipase (PLC-γ1); which in turn activate their downstream effectors
[30], including stimulation of the mitogenic protein kinase (MAP) cascade and protein kinase B (Akt). Afterward, diacylglycerol (DAG) and inositol triphosphate (IP3) are produced by IP3K activity and PLCγ1, leading to calcium (Ca
2+) mobilization. These pathways activate transcription factors involved in cell differentiation, survival, growth, and apoptosis, all of which are processes that occur in hours or days. In addition, NTFs modulate plasma membrane receptors, such as NMDA which are ionic channels permeable to Ca
2+ and Na
+; both Ca
2+ and Na
+ are crucial for neuronal function and differentiation
[58][59][59,60].
Melatonin receptor 1 (MT
1) and melatonin receptor 2 (MT
2) share amino acid sequences. However, other sequences distinguish them as “fingerprints”. MT
1 and MT
2 receptors are coupled to Galpha proteins which are stimulatory (Gs) or inhibitory (Gi). MT
1 and MT
2 can be coupled to Gi which in turn inhibits the cAMP production or to a Gq which activates beta-type PLC-β leading to the production of PIP2, IP3, and DAG
[53][60][54,61]. Similarly, to NTFs that activate TrKs receptors, MEL activates the same signaling pathways but through its binding with its specific receptors. In addition, MEL can cross the plasmatic membranes thanks to its amphiphilic features and binds to intracellular proteins such as calmodulin (CaM), calreticulin, and PKC, which transduce the biological responses triggered by Ca
2+. Furthermore, MEL binds to the quinone reductase-2 (the MT3 receptor) in the cytosol and the orphan retinoid receptors in the nucleus
[60][61][61,62].
Finally, it is worth mentioning that the Jun-kinase pathway is one of the most important pathways activated by NTFs. The signaling of NFTs involves protein 53 (p53) activation and apoptosis. Among the various targets of p53 is the proapoptotic gene Bax
[62][73]. Activation of one of the NTFs receptor p75NTR can also control the activity of Rho GTPase proteins, resulting in the inhibition of axonal growth and thus leading to selective pruning in neurodevelopment. In contrast, MEL modulates the apoptotic process via COX-2, p300, and the nuclear factor kappa β (NF-κβ) signaling, and, in addition, it suppresses p300 histone acetyltransferase (HAT) activity and p300-mediated NF-κβ acetylation
[63][74]. The information recapitulated shows that the effects of MEL are comparable to other NTFs’ functions, such as the proliferation of neuronal precursors, their survival, and their consequent differentiation between neurons or glial cells. Both Mel and NTFs share IP3 and Ca
2+-CaM signaling pathways.
As
we ha
s beenve described, the regulation of neurodevelopment through mechanisms elicited by NTFs allows to claim that the use of these molecules could be beneficial in the treatment of neuropsychiatric diseases. Moreover, neurogenesis studies have been a valuable tool for the development of new therapeutical drugs for neurodegenerative and affective disorders. In this context, it is important to mention that most antidepressants cause the release of NTFs that in turn stimulate neurogenesis in the hippocampus
[64][75], synaptogenesis in the prefrontal cortex
[65][76], and the production of neurogenic transcription factors
. [26][27][29][57][66][67][68][26,27,29,58,77–79].
In this regard, evidence accumulated in the last decade shows that MEL antidepressant-like effects in rodents are associated with increased neurogenesis in the dentate gyrus of the hippocampus [11][69][70][71][11,80–82]. Administration of MEL reduced the immobility time in the forced swimming test (FST) and in the tail suspension test (TST) paradigms, providing evidence for the antidepressant effects of MEL [72][73][83,84]. In this respect, one must add that the effect of MEL was potentiated depending on the time of administration [71][82].
In summary, the researchers collected evidence indicating that MEL is a factor that modulates neuronal survival, proliferation, differentiation, apoptosis, and the structural polarization of neurons. Remarkably, NTFs and MEL act similarly although they are chemically distinct. The former are peptides in nature while the latter is an indolamine with a low molecular mass. In addition, MEL is endogenously produced like neurotrophins and its actions can be mediated by specific receptors that activate signaling pathways promoting these processes. In addition, and similarly to NTFs, MEL stimulates neurogenesis in organisms at the early stages of their development and in the adult brain, promoting dendritogenesis and the extension of dendritic trees, particularly in the hippocampus.In summary, we collected evidence indicating that MEL is a factor that modulates neuronal survival, proliferation, differentiation, apoptosis, and the structural polarization of neurons. Remarkably, NTFs and MEL act similarly although they are chemically distinct. The former are peptides in nature while the latter is an indolamine with a low molecular mass. In addition, MEL is endogenously produced like neurotrophins and its actions can be mediated by specific receptors that activate signaling pathways promoting these processes. In addition, and similarly to NTFs, MEL stimulates neurogenesis in organisms at the early stages of their development and in the adult brain, promoting dendritogenesis and the extension of dendritic trees, particularly in the hippocampus.
Despite considerable efforts made in this field by specialists, MEL’s mechanisms of action have not been thoroughly understood and more knowledge gaps arise when it is used in combination with other substances. For this reason, it is necessary to conduct more research at molecular level to determine, firstly, whether these types of interactions exist, secondly, to understand these mechanisms in depth together with the signaling pathways that are activated to conduct their functions, and thirdly to study the utility of melatonin in the treatment of neurodegerative and neuropsychiatric diseses.