Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Masaki Arata and Version 2 by Conner Chen.
As exemplified by the unidirectionally beating cilia of multi-ciliated cells, various epithelial cells polarize not only along the apical-basal axis (inside–outside axis) of epithelial tissues, but also on the plane of epithelial tissues. The latter cell polarity, which is perpendicular to the apical–basal axis, is referred to as planar cell polarity (PCP). Pioneering research using the wings of Drosophila melanogaster identified a group of proteins, core PCP proteins, that orchestrate the establishment of PCP.
- cytoskeleton
- motile cilia
- multi-ciliated cells
- planar cell polarity
1. Introduction
Multi-ciliated cells line the surface of the oviduct, the trachea, and the ventricle of the brain (Figure 1A,B show oviduct multi-ciliated cells). At the apical surface of multi-ciliated cells, tens to hundreds of motile cilia are formed. The directions of cilia movements are unidirectionally aligned within each cell (rotational polarity), and they are consistent with the orientation of the tissue axis (tissue-level polarity). In addition, cilia exhibit a metachronal wave, a wave-like propagation of cilia movements in the plane of the tissue, which is produced by a temporal coordination of cilia movements between neighboring cilia (Figure 1C–D’) [1][2][3][4][5][1,2,3,4,5]. The coordination of cilia movements is essential for the functions of organs. In the mammalian oviduct, cilia pick up ovulated oocytes at the ovary end of the oviduct and carry them to the uterus [6][7][6,7]; in the trachea, cilia transport mucus and eliminate debris and pathogens [8][9][8,9]; and in the ventricle, cilia generate a flow of cerebrospinal fluid that is required for homeostasis of the organ [10].
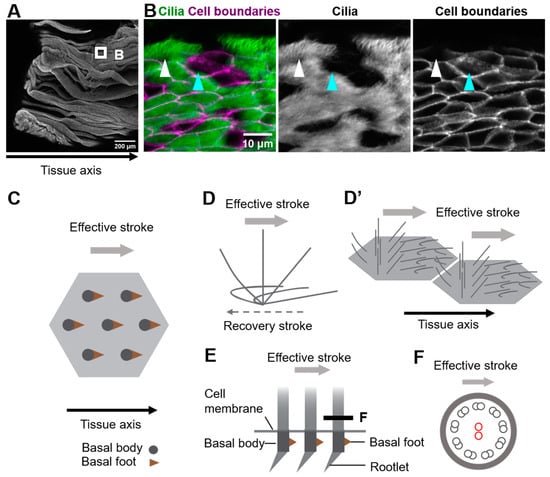
Figure 1. Coordination of cilia movements in multi-ciliated cells. (A) Ovary end of the oviduct was opened longitudinally and stained with E-cadherin. (B) Cilia (green; acetylated tubulin) and cell boundaries (magenta; E-cadherin) of the oviduct epithelium are visualized. The oviduct epithelium is composed of multi-ciliated cells (white arrowhead) and secretory cells (cyan arrowhead). At the apical surface of those multi-ciliated cells, about 150 cilia are formed on average. (C) Representative apical views of multi-ciliated cells. Gray circles and brown triangles indicate basal bodies and basal feet, respectively (also see Figure 1(E)). Note that the basal foot points in the same direction as the effective stroke (gray arrow). (D) Schematic representation of the movement of an individual cilium. Cilia repeat cyclic movements comprised of a fast effective stroke (gray arrow), and a slow recovery stroke (a backward motion; dotted arrow). (D’) A schematic of cilia movements in multi-ciliated cells. Apical surfaces of two multi-ciliated cells are shown. Note that the orientation of effective stroke (or recovery stroke) is consistent with that of the tissue axis. The phase of the beating cycles of cilia shifts between neighboring cilia and generates a metachronal wave, which is a wave-like propagation of cilia movements. (E) Lateral view of the basal region of cilia. The basal body has two appendages, the basal foot and the rootlet. (F) Cross-sectional view of the cilium along the gray line in (E). 9 + 2 microtubules are shown in small circles. The central pair of 9 + 2 microtubules (red circles) run perpendicular to the direction of the effective stroke (gray arrow).
Motile cilia of multi-ciliated cells are microtubule-based cell protrusions which show a biphasic movement comprised of a fast effective stroke, and a slow backward motion, the recovery stroke (Figure 1D). Inside the majority of motile cilia, 9 + 2 arrays of microtubules run along the longitudinal axis of the cilium, and a central pair of microtubules lie perpendicular to the beating orientation of the cilium (Figure 1E,F) [11]. The basal body lies at the base of cilium, and the basal foot and the rootlet are associated with the basal body (Figure 1E). A single basal foot protrudes from the lateral side of each basal body, and its direction is consistent with that of the effective stroke [12][13][12,13]. A rootlet is located at the proximal end of each basal body and extends to the center of the cell [14][15][16][17][14,15,16,17].
2. Roles of Core PCP Proteins in Coordinating Cilia Orientation
As exemplified by the unidirectionally beating cilia of multi-ciliated cells, various epithelial cells polarize not only along the apical-basal axis (inside–outside axis) of epithelial tissues, but also on the plane of epithelial tissues. The latter cell polarity, which is perpendicular to the apical–basal axis, is referred to as planar cell polarity (PCP) [18][19][20][21][22][23][24][25][26][27][28][18,19,20,21,22,23,24,25,26,27,28]. Pioneering research using the wings of Drosophila melanogaster identified a group of proteins, core PCP proteins, that orchestrate the establishment of PCP (Figure 2). Core PCP proteins are an evolutionally conserved group of proteins comprised of transmembrane proteins, Flamingo/Starry night (Fmi/Stan), Van Gogh (Vang) and Frizzled (Fz), as well as cytoplasmic proteins Prickle (Pk), Dishevelled (Dvl) and Diego (Dgo) [21][26][29][30][21,26,29,30]. Each Drosophila wing epithelial cell forms an actin-rich cell protrusion, a wing hair, at the apical cortex and each wing hair points to the distal end of the wing, which is a hallmark of PCP in the wing [21]. Just before the onset of wing hair formation, cell boundary localization of core PCP proteins is strongly biased along the tissue axis (Figure 2A). Fz- and Vang-containing complexes (referred to as the Fz- and Vang-complex in Drosophila, respectively, or the FZD- and VANGL-complex in vertebrates, as used hereafter) localize at the distal and proximal side of the cell, respectively (Figure 2A,A’) [28][31][32][33][34][35][28,31,32,33,34,35]. When each member of core PCP proteins is lacking, the orientations of wing hairs are not coordinated along the tissue axis [21]. In addition to the Drosophila wing, the asymmetric distribution of core PCP proteins was observed in various organs and animals, including multi-ciliated cells of the mouse oviduct, trachea, and ventricle (Figure 2B) [20][36][37][38][20,36,37,38]. The establishment of the polarized distribution of core PCP proteins precedes the formation of multi-cilia in the developing oviduct [20][39][20,39]. Furthermore, the loss of core PCP proteins abrogates the orientation of cilia, suggesting that the asymmetric distribution of core PCP proteins provides a cue to orient cilia. In contrast to Drosophila genes encoding core PCP proteins, those in vertebrates are often duplicated and might have divergent functions (Figure 2A”). For example, there are three homologs of Drosophila Flamingo, CELSR1, 2, and 3 in mice, and these have different expression patterns and show different phenotypes when their functions are lost [20][40][41][42][20,40,41,42].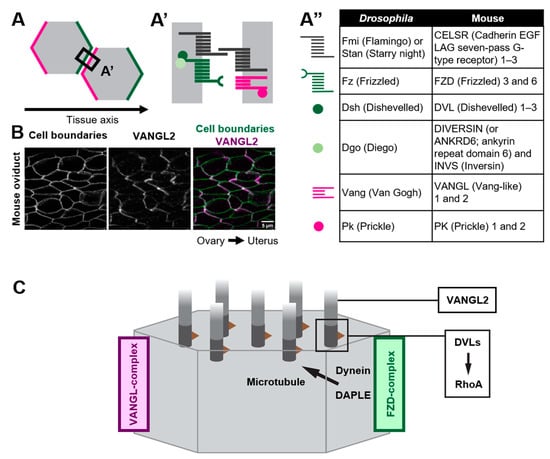
Figure 2. Polarized distribution of core PCP proteins along the tissue axis. (A–A”) Core PCP proteins form an asymmetric complex at cell boundaries. (A,A’) FZD-containing complex (FZD-complex; green) and VANGL-containing complex (VANGL-complex; magenta) are segregated to opposite cell boundaries (note that their distribution is polarized along the tissue axis). Extracellular domain of CELSRs provides intercellular bridges between the FZD-complex and the VANGL-complex, thus enabling the coupling of cell polarity at a multicellular level. (A”) A list of members of core PCP proteins in Drosophila and their counterparts in mice. (B) Mouse oviduct epithelium was stained for E-cadherin and a core PCP protein, VANGL2. E-cadherin labels cell boundaries. Note the zigzag pattern of VANGL2 signals which highlights polarity in the cell-boundary distribution of VANGL2 along the ovary–uterus axis. (C) Mechanisms by which core PCP proteins control the coordinated movements of cilia.