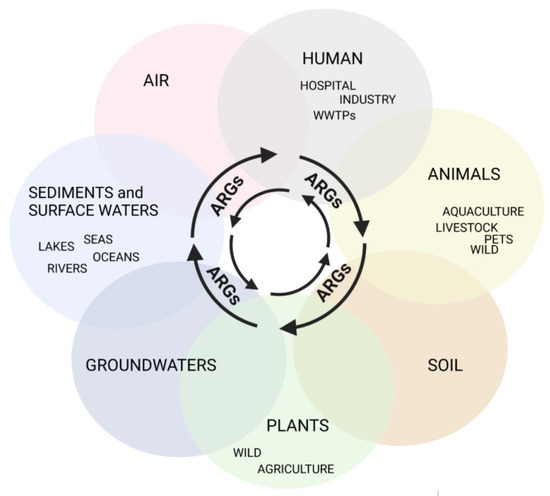
Antimicrobial resistance (AMR) is one of the largest global concerns due to its influence in multiple areas, which is consistent with One Health’s concept of close interconnections between people, animals, plants, and their shared environments. Antibiotic-resistant bacteria (ARB) and antibiotic-resistance genes (ARGs) circulate constantly in various niches, sediments, water sources, soil, and wastes of the animal and plant sectors, and is linked to human activities. Sewage of different origins gets to the wastewater treatment plants (WWTPs), where ARB and ARG removal efficiency is still insufficient, leading to their transmission to discharge points and further dissemination. Thus, WWTPs are believed to be reservoirs of ARGs and the source of spreading AMR. According to a World Health Organization report, the most critical pathogens for public health include Gram-negative bacteria resistant to third-generation cephalosporins and carbapenems (last-choice drugs), which represent β-lactams, the most widely used antibiotics. Therefore, this research presents the available research data for ARGs in WWTPs that confer resistance to β-lactam antibiotics, with a particular emphasis on clinically important life-threatening mechanisms of resistance, including extended-spectrum β-lactamases (ESBLs) and carbapenemases (KPC, NDM).
- antibiotic resistance genes
- wastewater treatment plant
- β-lactamase
- extended-spectrum β-lactamase
- carbapenemases
1. Introduction
2. Clinically Significant β-Lactam Resistance Genes in Wastewater Treatment Plants—The Occurrence and Distribution
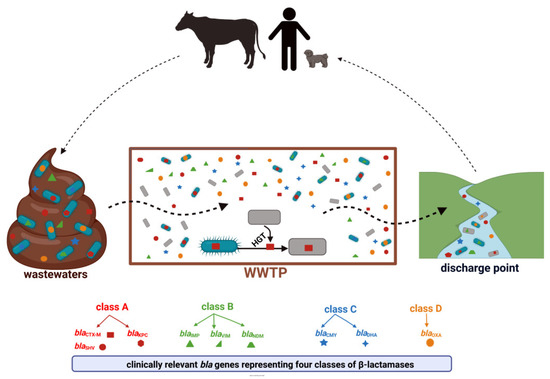
2.1. Class A β-Lactamases
2.1.1. Class A β-Lactamases—Occurrence and Variability in WWTPs-Linked Samples
Due to the global spread of class A β-lactamases, it is a commonly, or even predominantly, detected group in WWTPs. In a multi-national study of WWTPs from Denmark, Spain, and the United Kingdom (UK) with high-throughput qPCR used, these β-lactamases was leading, accounting for 70% of all detected bla genes [[652]]. Among them, the most relevant were two groups linked with ESBL and KPC enzymes. It is noteworthy that, among the ESBL group, the most common in clinical settings and in various wastewater sources is CTX-M encoded by blaCTX-M, carried mainly by Enterobacterales [53][66]. In this review, blaCTX-M was detected in the majority of included studies and, in many, had the highest prevalence [54][55][5[67,68,6][59,7][58][59][60][61][62][0,71,72,73,74,75,763][64][65][66][67][6,77,78,79,80,8]1]. However, blaSHV and/or blaTEM were found frequently as well [15][69][70][71][72],82,83,84,85]. In some studies, blaTEM was predominant, e.g., in an Irish study [73][86], as well as in Colombia [[874]], Poland and Portugal [9][75],88], Belgium [76][89], the US [77][90], and Africa [78][91]. Another significant group representing the KPC family encoded by blaKPC genes was detected in numerous WWTPs from European [[652][5,6][76][79,89,9][80][81][82][83][84][2,93,94,95,96,97,985][86][87][88],99,100,101], as well as from American [77][89][9[90,102,10][93,1]04], African [59][78][92][72,91,105], and Asian countries [93][94][106,107]. Moreover, analysis of reviewed articles, especially those using developed techniques as high-throughput qPCR, whole-genome sequencing, or metagenomics, shows a high variety of detected genes of the discussed β-lactamases, not only representing blaCTX-M, blaSHV, blaTEM, and blaKPC families, but also others less frequently associated with public health, i.e., BEL, cfxA, GES, PER, SME, VEB, and others [[652][7,9][83][89][94][95][2,96,102,107,108,1096][97][98],110,111]. Environmental studies based on the analysis of bacterial strains during the treatment process most often concern the most critical pathogens posing the greatest threat, mainly Enterobacterales. In the reviewed literature, the predominantly tested and detected species among this bacteria family were Escherichia coli and K. pneumoniae [54][55][58][59][61][64][65][66][6[67,68,71,72,74,77,78,79,8][70][72][86][99][1,83,85,99,112,1100][3,101][4,1102][5,1103][6,1104][7,1105][8,1106][9,1207][,12108][,109][22,110][23,111][24,112]5]; however, different species of Citrobacter spp., Enterobacter spp., Pseudomonas spp., Aeromonas spp., or others were noted as well [58][[71,763][67][,80,874][7,9][89][92,102,1][04,1096][97][,110,12108][,113][26,114][27,115]28]. It is noteworthy that antibiotic susceptibility testing of the studied ESBL-producing strains isolated from the WWTPs confirms a high percentage of multi-drug resistance. It was also noted that these bacteria may survive the treatment process and that the WWTPs were unable to eradicate them completely. Generally, the number of MDR isolates decreased during the treatment, but for some, their proportion was still significant in effluents, in some even higher than in influent samples [5[7][58][60][73][75][70,71,73,86,88,9][81][84][86][2,94,97,99,1105][8,1106][9,110][123,12][5,116][29,117]30]. Moreover, analyzing downstream river or marine samples where final effluents are released, MDR isolates carrying ESBL enzymes were commonly detected [20][66][75][7,79,88,9][2,117]30]. Molecular typing concerning bacteria isolated from WWTPs confirmed high genetic relatedness between bacteria from WWTPs and human- and animal-associated sources, as well as the presence of clinically important lineages such as pandemic ST131 E. coli in WWTPs-related samples. Liedhegner et al. compared E. coli isolated from samples of various environmental compartments from one geographic area (clinical samples, hospital wastewater, and WWTP). The data including antibiotic resistance, virulence, and ESBL gene profiles confirmed high phenotypic and genotypic similarity across strains of these different origins and demonstrated potential health risks related to ESBL transmission [1[12]5]. An interesting study conducted by Raven et al. showed genetic relatedness between E. coli isolated from 20 WWTPs in the UK, livestock farms, retail meat, and isolates responsible for human blood infections. The genomic analysis of i.e., ESBL-producing isolates revealed that the three most common sequence types (STs) associated with bloodstream infections (ST131, ST73, and ST95) and the specific and most common for livestock (ST10) were found in wastewater samples [1207]]. In many other studies, human-associated, multidrug-resistant, and highly virulent clone ST131 E. coli was detected in WWTP samples as well [62][[75,874][,1100][3,1318][,119][32,120][33,121]34].2.1.2. Class A β-Lactamases—Removal during the Treatment Process
Concerning the removal of class A β-lactam ARGs, there is no universal target panel in qPCR studies; however, it has been noted that, although the WWTPs could effectively eliminate examined genes, their abundance was still reported in effluents and receiving water bodies. For example, in the study of Schages et al., strains harboring blaCTX-M were isolated from the effluent [110]23], as well as in a Japanese study wherein strains possessing ARGs belonging to the blaCTX-M-1, blaCTX-M-9, blaTEM, and blaSHV families survived even after sterilization [111]24]. Other studies reported similar results of the ARGs’ presence in effluent samples [95][[108,122][35,123][6,124][37,125]38]. In Polish research from Kozieglowy, it was noticed that the wastewater treatment process leads to a significant increase in the relative abundance of blaTEM and blaGES genes, while the abundance of blaKPC decreased. Finally, the removal efficiency of ARGs was the least for blaGES (94.8%) and blaCTX-M (95.3%), while for other genes, it was >98% [5[6]9]. In another study, the presence of blaKPC was completely eliminated even after the first mechanical procedure [80][93]. In a Chinese survey comparing bacteria carrying blaCTX-M, blaSHV, and blaTEM, isolated from influent and effluent, higher prevalence was noted in influent samples, except for blaCTX-M, which was more frequently detected in effluent samples [116]29]. Significant differences between influent and effluent were described in a Romanian investigation and concerned blaSHV-100, -145, which were decreased during treatment [72][85]. Interestingly, Neudorf et al. analyzed 3 WWTPs in Arctic Canada and noted a decrease of blaTEM abundance in two sites with a passive system and no significant changes for a third WWTP with a mechanical system. Moreover, no differences were found for blaCTX-M in all treatment plants [126]39]. A Spanish study by Rodriguez-Mozaz et al. demonstrated an increased frequency of blaTEM during the treatment process [127]40], while in a study of three WWTPs from Finland and Estonia, no significant changes were noted for blaCTX-M-32, unlike blaSHV-34, of which the relative concentration was increased in effluent samples but only in one tested WWTP [[14128]]. Comparable data with similar blaCTX-M and blaTEM concentrations in influent and effluent samples were obtained in a study of five WWTPs in Tunisia; however, the abundance of the genes was higher in the effluent in a WWTP receiving additional hospital wastewater [1429]]. The occurrence of class A β-lactamases ARGs was also detected in downstream river samples whence final effluents were discharged, e.g., in a multi-national study including sixteen WWTPs from ten European countries [88][101], in a study conducted by Zieliński et al., wherein the predominant blaTEM was noted in receiving river water samples [15], and in a study performed by Osińska et al., wherein the presence of blaSHV and blaTEM in receiving river samples was confirmed [71][84]. WWTPs pose a health risk, not only because treated wastewater containing AMR genes or MDR bacteria are transferred into surface water bodies, but also because these pollutants are discharged into the air surrounding WWTPs through bioaerosol generated from bioreactors [15][55],68]. The study of the carriage of ESBL-producing Enterobacterales in WWTP workers and surrounding residents shows that these groups are much more like to acquire bacteria harboring the ESBL mechanism [25], thus confirming the direct influence of WWTPs on spreading ARGs into air. The contribution of WWTPs’ bioaerosols in ARGs and ARB propagation into air and different environments is commonly investigated [1430][,131][44,132][45,133]46]. For example, Gaviria-Figueroa et al. studied bioaerosol samples collected downwind from sludge aeration tanks and showed a significant presence of clinically relevant class A β-lactamases, along with other classes of these enzymes and different antibiotic groups [134]7].2.2. Class B β-Lactamases
2.2.1. IMP and VIM β-Lactamases in WWTPs-Linked Samples
As with the previously discussed ARGs, the environment plays a role in the transmission of blaIMP and blaVIM encoding MBLs enzymes with clinical importance, IMP and VIM, respectively. Although the majority of reports focus on hospital wastewater, these genes were detected also in samples of wastewater treatment plants from the US [69][89][9[82,102,10][3,134]7], Canada [9[1]04], China [5[7][69]0,82], and Singapore [94][107] as well as from many European countries, such as Sweden [83][[96,1096]], Switzerland [86][99], the UK [115]28], Germany [87][[100,110][23,123][6,150]62], Poland [5[6][79,9][80][2,93,151]63], Slovakia [[1102]5], and Romania [81][94]. A multi-national study concerning urban WWTPs in Denmark, Spain, and the UK showed the permanent presence of blaVIM during the treatment process even in downstream river samples, in contrast to other tested genes, which were reduced under a detectable level [[652]]. Interesting results were presented by Khan et al., who compared Klebsiella oxytoca strains isolated from clinical sources (hospital wastewater) and the river receiving effluents from WWTP in Örebro, Sweden. Results obtained for two selected strains—the same antibiotic susceptibility patterns, antibiotic resistance gene profiles (i.e., blaVIM-1, blaOXA-10, blaACC-1), MLST type, furthermore phylogenetic relationship based on core genome single nucleotide polymorphism (SNP) analysis, and core genome MLST—suggest the transfer of K. oxytoca-producing carbapenemases from the hospital setting to the aquatic environment, which may pose a threat to the community [152]64].2.2.2. New Delhi Metallo-β-Lactamase (NDM) in WWTPs-Linked Samples
According to epidemiological data, NDMs seem to pose the greatest threat among class B β-lactamases. Genes encoding them were noted in many aquatic environments, including animal production wastewaters, industrial, domestic sewage, tap water, surface water, and groundwater. However, hospital wastewater is considered to be a major source of blaNDM variants [1653][,154][66,155]67]. As the geographical origin of NDM-producing bacteria is India, multiple publications detecting blaNDM, especially in hospital sewage, come from India [156][8,157][69,158]70], together with other Asian [95][[108,17159][,160][72,161]73] and African countries [92][[105,162]74]. Interesting results were reported by Marathe et al., who studied hospital wastewater from Mumbai, India. Shotgun metagenomics revealed the presence of β-lactamase genes encoding clinically important MBLs, such as NDM, VIM, and IMP with blaNDM as the most common carbapenemase-encoding gene. Additionally, 27 unique MBL genes not known yet were detected, which showed the huge potential of the metagenomic approach [163]75]. However, NDM-lactamases in Asian countries were not only detected in hospital sewage samples. Analysis of rivers and sewage treatment plants in five Indian states also showed an abundance of blaNDM [64][77]. Similarly obtained data from southwest China showed a wide distribution of blaNDM in hospital sewage, WWTP effluent, and river samples. Interestingly, the gene was found in many different bacterial species belonging to Enterobacterales, genus Acinetobacter, and Pseudomonas [1764]]. The data from northern China [165][77,166]78] and Saudi Arabia [167]9] also confirm the presence of blaNDM in WWTP samples. blaNDM has spread globally, and several variants were noted not only in India and China but in many other countries in various water samples, including those from WWTPs and the surface waters of WWTP discharge points in the UK [115]28], Belgium [76][89], Switzerland [86][99], Germany [87][100], Poland [5[6][9,151]63], the Czech Republic [168]0], Romania [72][81][85,94], Spain [[985]], Africa [78][92][91,105], and the US [77][89][9[90,102,10]3]. Interesting results concern the Irish study conducted by Mahon et al. They examined the genetic relationship between NDM-possessing E. coli and K. pneumoniae (separately) cultivated from three locally linked sources: sewage samples from the collection system, freshwater streams, and clinical isolates. E. coli were considered indistinguishable, and K. pneumoniae were very closely related. These results confirm that water sewage plays an important role in the resistance transfer process [[18169]]. Another analysis by Walsh et al. concerning public tap water and seepage water from sites around New Delhi also indicates that the environment has an undeniable influence on the propagation of NDM resistance [170]82]. Data regarding the wastewater treatment process show a different level of the transmission of bacteria with the NDM mechanism during the treatment process and the effectiveness of blaNDM reduction. In a Polish urban WWTP from Kozieglowy, Makowska et al. studied β-lactamase genes in the genomes of ESBL-producing and carbapenem-resistant coliforms isolated from each stage of the treatment process. They found that blaNDM and blaVIM were present in all stages and that the highest frequency was recorded in isolates from effluent compared to raw sewage, which indicates that the treatment process in the mechanical–biological treatment plant is insufficient in eliminating blaNDM and the organisms carrying them [5[6]9]. Similarly, data from two WWTPs in north China show the persistent and prevailing presence of blaNDM even after disinfection [165]77] and the propagation of blaNDM from a WWTP into its receiving river [166]78]. Other studies measuring absolute (copies/mL) and relative (copies/16S) abundance of blaNDM in influent and effluent also confirm deficient reduction [[985][,171]83]. However, Divyashree et al., who studied treated and untreated effluents from hospital samples in Mangalore, South India, showed the absence of blaNDM in treated effluents [172]84]. A Polish study also showed a complete reduction of blaNDM in the treatment process, even after the initial treatment stage [80][93], similar to a multi-center study from Denmark, Spain, and the UK [[652]].2.3. Class C β-Lactamases
2.3.1. Class C β-Lactamases in WWTPs-Linked Samples
2.4. Class D β-Lactamases
2.4.1. OXA Family β-Lactamases Carried in ARB
The reviewed approaches concerning class D β-lactamases are focused on bacterial strains carrying blaOXA isolated from WWTP samples. The majority of these studies confirm a blaOXA presence in isolates from both untreated and treated samples, and the prevalent variants are blaOXA-1 and blaOXA-48. Multiple examples come from European countries: a Czech study reported ESBL-producing Enterobacterales carrying blaOXA-1 and isolated from effluent; globally spread MDR clones of E. coli ST131 and K. pneumoniae ST321 and ST323 harboring large FIIK plasmids with multiple antibiotic-resistance genes were found among tested strains [[1100]3]; a Spanish study detected blaOXA-1 in strains isolated from effluents of two out of 21 tested WWTPs [[763]]; two German studies reported the presence of blaOXA-51 and blaOXA-48 in carbapenemase-producing bacteria [87][100] and blaOXA-58, blaOXA-48 and blaOXA-23 in bacterial strains isolated from influent, activated sludge and effluent [110]23]; four Polish studies identified blaOXA genes among ceftazidime- or meropenem-resistant bacterial strains [7[9]2], Aeromonas spp. strains isolated from raw sewage, activated sludge, and effluent [84][97], ESBL-producing Enterobacterales [55][68] and Acinetobacter spp. isolates [151]63]; an Austrian study of carbapenemase-producing Enterobacterales from activated sludge confirmed harboring blaOXA-48 [82][95]; and a study concerning the WWTP in Basel, Switzerland, where carbapenemase-resistant Enterobacterales and other Gram-negative bacteria isolated from municipal and hospital wastewater and WWTP receiving this sewage were compared, and identical isolates from the WWTP and wastewater samples were detected, including OXA-48-producing E. coli ST38 and Citrobacter spp. [86][99]. Similarly, a molecular epidemiology approach was conducted in a Romanian study. Surleac et al. detected variants of blaOXA in K. pneumoniae isolated from samples of three WWTPs [72][85], while Teban-Man et al. compared carbapenemase-producing K. pneumoniae isolated from the influent and effluent of two WWTPs with and without hospital input and found that blaOXA-48 was carried by strains isolated from raw and treated samples of WWTPs collecting hospital wastewater. In the second WWTP, the gene was observed only in strains from influent. Moreover, isolates harboring blaOXA-48 were genetically typed, which showed they belonged to sequence types of high-risk clones (ST258, ST101, ST147, ST2502). These clones were associated with clinical settings and reported to be multi-drug resistant [81][94]. In a study of a Swedish WWTP, Gram-negative bacteria harboring blaOXA were noted in influent, effluent, and recipient waters of the river and lake [[1096]]. However, in a Portuguese study conducted by Araujo et al., blaOXA was detected only in strains isolated from raw sewage samples [97][110]. Another Portuguese investigation of ampicillin-resistant Enterobacterales isolated from influent and effluent showed different results; blaOXA was the most prevalent gene among tested ESBL-producing strains [75][88]. There are significantly fewer studies detecting blaOXA in the African region and they cover Algeria, where blaOXA-1 was detected [61][74]; Durban, South Africa, where cefotaxime-resistant E. coli were studied and blaOXA-1 was found as well [78][91]; Eastern Cape Province, South Africa, where blaOXA-1-like and blaOXA-48-like variants harbored by Enterobacterales isolated from effluents of WWTPs were noted [59][72]; and Tunisia, where C. freundii isolate carrying blaOXA-204 [[12108]] and Enterobacterales strains possessing blaOXA-1 [114]27] were detected. American studies concerning WWTPs also confirm blaOXA presence in bacteria isolated from WWTP samples [67][77][89][9[80,90,102,1][04,1106][9,112][5,120]33].2.4.2. OXA Family β-Lactamases in Direct WWTP Samples—Occurrence and Removal
Multiple studies report the presence of blaOXA in direct WWTP samples and determine the concentration and relative abundance of selected gene variants to define the efficiency of the treatment process. Comparable to previously discussed β-lactamases, bacteria producing OXA enzymes, as well as blaOXA, can be detected after the treatment process. For example, the study of two WWTPs in the Brussels region determined the relative abundance of blaOXA-48 in different stages of the treatment, as well as in samples of the river as the discharge point for the WWTP effluents. In that study, Proia et al. showed a significant increase of blaOXA-48 from influent to effluent and from upstream to downstream river samples [76][89]. Similarly, in Kozieglowy, a Polish WWTP, it was reported that the wastewater treatment process leads to a significant increase in the relative abundance of blaOXA-48 genes in the effluent [5[6]9], whereas in research from the Baltic Sea area, the relative abundance of blaOXA-58 was decreased in the effluent; however it was weakly significant and found only in one of the three studied WWTPs [[14128]]. In the German study, the absolute abundance of selected blaOXA genes was determined, and when comparing raw and treated samples from WWTP, a significant decrease was reported regarding blaOXA-58 and blaOXA-48 but not blaOXA-23 [110]23]. Similar results were obtained in a multi-national study of WWTPs from ten European countries, where qPCR and absolute abundance were performed for selected blaOXA genes. It was noticeable that, among all tested β-lactamase genes, blaOXA-58 was found in all tested samples, had the highest absolute abundance, and was significantly reduced during treatment [88][101]. In three Swedish municipal sludge treatment plants, a metagenomics approach was conducted, and many variants of blaOXA were detected at all stages of the treatment process. Some of them, like blaOXA-48, were consistently enriched in treated sludge compared to primary sludge [83][96]. Other metagenomic approaches or using qPCR provide similar results—the presence of multiple blaOXA gene variants, including effluent samples [80][[93,1096][,123][6,134]7], while others detected only single or a few variants [[652][5,6][76][88][94][95][98][9,89,101,107,108,111,14128][,151]63]. Interestingly, in a Polish study, where blaOXA was detected as one of the prevalent tested genes in influent and effluent samples, comparative metagenomic analysis of DNA from WWTP samples and employees’ swabs revealed the presence of similar ARGs in both types of samples with significantly higher concentrations than in control samples [15]. Other studies that report the presence of blaOXA genes at different stages of the treatment process include the research of Yang et al., wherein activated sewage sludge from 15 WWTPs was tested, and three variants (blaOXA-1, blaOXA-2 and blaOXA-10) were detected [69][82], while in WWTP active sludge in South Carolina, in the US, a higher variability among blaOXA (seven variants) was noted [134]7]. Interesting results concerning the seasonal increase of blaOXA concentration between the summer and winter seasons were reported in the study of four small-scale domestic WWTPs. Furthermore, blaOXA in winter was prevalent among tested ARGs in raw sewage, as well as in effluent samples; additionally, the gene was detected in receiving river samples, in both the winter and summer seasons [71][84]. Results of a multi-national study, analyzing samples from Denmark, Spain, and the UK, indicated a country-specific presence for blaOXA-10 detected only in WWTPs from the UK [[652]]. The above data, showing the presence of blaOXA genes and bacteria harboring them in WWTPs and related samples, confirms that WWTPs are a hotspot for antibiotic-resistant gene transmission into not only the aquatic compartments of the environment but also to the atmospheric air, creating an additional health risk for the workers of WWTPs.3. Conclusions
AMR is a serious and urgent problem, and it is clear that the environment plays a key role in the process of transmission and propagation of ARGs and ARB with life-threatening clinical consequences. The multitude of publications confirms that β-lactamases genes encoding especially ESBLs (TEM, SHV, CTX-M) and KPC, NDM, and OXA carbapenemases, which pose one of the greatest health risks, are widely found in WWTPs and disseminated to further portions of the environment. Molecular analysis shows repeatedly high genetic relatedness between environmental and clinical isolates, e.g., ST131 E. coli. Generally, different kinds of sewage treatment processes do not eliminate these ARGs completely. Furthermore, some data indicate an increased level of β-lactam ARGs in effluent or even the presence of the genes and bacteria harboring them in samples after additional disinfection treatments. Due to β-lactam ARGs’ potential to transfer via mobile genetic elements through horizontal gene transfer, their abundance in water samples discharged from WWTPs into natural aquatic sources used by humans or animals suggests a potential risk of transmission resistance determinants into pathogenic and non-pathogenic bacteria and acquiring multidrug resistance as well as the participation of WWTPs in AMR transmission route and distribution into surrounding ecosystems and clinical settings. The growing problem of AMR and the spread of clinically relevant ARGs related to, i.e., β-lactams in the environment, indicate the need to improve and evaluate the procedures of wastewater treatment and disinfection; thus, ARB, ARGs, and factors influencing their selection and co-selection during the treatment process would be completely removed. The development and improvement of techniques used in testing wastewater for antibiotic resistance has been very significant in recent years. There are more and more publications indicating the use of modern metagenomic assays, which enables broadening the knowledge of the complexity and structural and functional biodiversity of microbial communities—i.e., analysis of resistance genes; taxonomic assignment; functional genes characterization; the identification of the HGT mechanism and mobile elements involved in the gene transmission; and exploring relationships between pathogenic and non-pathogenic species and susceptible and resistant bacteria. Therefore metagenomic analysis seems to be a very useful tool to understand the process of AMR transmission. However, the clinical surveillance of resistant strains responsible for life-threatening infections and nosocomial outbreaks caused by β-lactam-resistant strains also involve molecular techniques, but still the gold standard are culture-based methods detecting the expression of genes and the resistance mechanism. Therefore, according to the One Health’s concept, collaborative approaches concerning AMR in the environment and clinical setting are indispensable and should combine new technology with standard microbiological methods. As WWTPs are the crucial points on the routes of ARB and ARGs’ spread, they should be deeply explored, which would help to understand the process and make it possible to introduce procedures to stop, or at least slow down, the spreading of antibiotic resistance.References
- WHO. Ten Threats to Global Health in 2019; World Health Organization: Geneva, Switzerland, 2019; pp. 1–18. Available online: https://www.who.int/news-room/spotlight/ten-threats-to-global-health-in-2019 (accessed on 10 May 2022).
- CDC. Antibiotic Resistance Threats in the United States 2019. Available online: https://www.cdc.gov/drugresistance/pdf/threats-report/2019-ar-threats-report-508.pdf (accessed on 1 April 2022).
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327.
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Robles Aguilar, G.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655.
- Abushaheen, M.A.; Muzaheed; Fatani, A.J.; Alosaimi, M.; Mansy, W.; George, M.; Acharya, S.; Rathod, S.; Divakar, D.D.; Jhugroo, C.; et al. Antimicrobial resistance, mechanisms and its clinical significance. Disease-a-Month 2020, 66, 100971.
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281.
- Aslam, B.; Khurshid, M.; Arshad, M.I.; Muzammil, S.; Rasool, M.; Yasmeen, N.; Shah, T.; Chaudhry, T.H.; Rasool, M.H.; Shahid, A.; et al. Antibiotic Resistance: One Health One World Outlook. Front. Cell. Infect. Microbiol. 2021, 11, 1–20.
- Alexander, J.; Hembach, N.; Schwartz, T. Evaluation of antibiotic resistance dissemination by wastewater treatment plant effluents with different catchment areas in Germany. Sci. Rep. 2020, 10, 8952.
- Vaz-Moreira, I.; Harnisz, M.; Abreu-Silva, J.; Rolbiecki, D.; Korzeniewska, E.; Luczkiewicz, A.; Manaia, C.M.; Plaza, G. Antibiotic resistance in wastewater, does the context matter? Poland and Portugal as a case study. Crit. Rev. Environ. Sci. Technol. 2022, 52, 4194–4216.
- Pal, C.; Bengtsson-Palme, J.; Kristiansson, E.; Larsson, D.G.J. Co-occurrence of resistance genes to antibiotics, biocides and metals reveals novel insights into their co-selection potential. BMC Genom. 2015, 16, 964.
- Pazda, M.; Kumirska, J.; Stepnowski, P.; Mulkiewicz, E. Antibiotic resistance genes identified in wastewater treatment plant systems—A review. Sci. Total Environ. 2019, 697, 134023.
- Wang, J.; Chu, L.; Wojnárovits, L.; Takács, E. Occurrence and fate of antibiotics, antibiotic resistant genes (ARGs) and antibiotic resistant bacteria (ARB) in municipal wastewater treatment plant: An overview. Sci. Total Environ. 2020, 744, 140997.
- Felis, E.; Kalka, J.; Sochacki, A.; Kowalska, K.; Bajkacz, S.; Harnisz, M.; Korzeniewska, E. Antimicrobial pharmaceuticals in the aquatic environment—Occurrence and environmental implications. Eur. J. Pharm. 2020, 866, 172813.
- Uluseker, C.; Kaster, K.M.; Thorsen, K.; Basiry, D.; Shobana, S.; Jain, M.; Kumar, G.; Kommedal, R.; Pala-Ozkok, I. A Review on Occurrence and Spread of Antibiotic Resistance in Wastewaters and in Wastewater Treatment Plants: Mechanisms and Perspectives. Front. Microbiol. 2021, 12, 717809.
- Zieliński, W.; Korzeniewska, E.; Harnisz, M.; Drzymała, J.; Felis, E.; Bajkacz, S. Wastewater treatment plants as a reservoir of integrase and antibiotic resistance genes—An epidemiological threat to workers and environment. Environ. Int. 2021, 156, 106641.
- Miłobedzka, A.; Ferreira, C.; Vaz-Moreira, I.; Calderón-Franco, D.; Gorecki, A.; Purkrtova, S.; Bartacek, J.; Dziewit, L.; Singleton, C.M.; Nielsen, P.H.; et al. Monitoring antibiotic resistance genes in wastewater environments: The challenges of filling a gap in the One-Health cycle. J. Hazard. Mater. 2022, 424, 127407.
- Thomas, C.M.; Nielsen, K.M. Mechanisms of, and Barriers to, Horizontal Gene Transfer between Bacteria. Nat. Rev. Microbiol. 2005, 3, 711–721.
- von Wintersdorff, C.J.H.; Penders, J.; van Niekerk, J.M.; Mills, N.D.; Majumder, S.; van Alphen, L.B.; Savelkoul, P.H.M.; Wolffs, P.F.G. Dissemination of Antimicrobial Resistance in Microbial Ecosystems through Horizontal Gene Transfer. Front. Microbiol. 2016, 7, 173.
- Czekalski, N.; Gascón Díez, E.; Bürgmann, H. Wastewater as a point source of antibiotic-resistance genes in the sediment of a freshwater lake. ISME J. 2014, 8, 1381–1390.
- Kotlarska, E.; Łuczkiewicz, A.; Pisowacka, M.; Burzyński, A. Antibiotic resistance and prevalence of class 1 and 2 integrons in Escherichia coli isolated from two wastewater treatment plants, and their receiving waters (Gulf of Gdansk, Baltic Sea, Poland). Environ. Sci. Pollut. Res. 2015, 22, 2018–2030.
- Chaturvedi, P.; Chaurasia, D.; Pandey, A.; Gupta, P. Co-occurrence of multidrug resistance, β-lactamase and plasmid mediated AmpC genes in bacteria isolated from river Ganga, northern India. Environ. Pollut. 2020, 267, 115502.
- Griffin, D.W.; Banks, K.; Gregg, K.; Shedler, S.; Walker, B.K. Antibiotic Resistance in Marine Microbial Communities Proximal to a Florida Sewage Outfall System. Antibiotics 2020, 9, 118.
- Kvesić, M.; Kalinić, H.; Dželalija, M.; Šamanić, I.; Andričević, R.; Maravić, A. Microbiome and antibiotic resistance profiling in submarine effluent-receiving coastal waters in Croatia. Environ. Pollut. 2022, 292, 118282.
- Sabri, N.A.; Schmitt, H.; Van Der Zaan, B.; Gerritsen, H.W.; Zuidema, T.; Rijnaarts, H.H.M.; Langenhoff, A.A.M. Prevalence of antibiotics and antibiotic resistance genes in a wastewater effluent-receiving river in the Netherlands. J. Environ. Chem. Eng. 2020, 8, 102245.
- Rodríguez-Molina, D.; Berglund, F.; Blaak, H.; Flach, C.-F.; Kemper, M.; Marutescu, L.; Gradisteanu, G.P.; Popa, M.; Spießberger, B.; Weinmann, T.; et al. Carriage of ESBL-producing Enterobacterales in wastewater treatment plant workers and surrounding residents—The AWARE Study. Eur. J. Clin. Microbiol. Infect. Dis. 2021.
- Leonard, A.F.; Morris, D.; Schmitt, H.; Gaze, W.H. Natural recreational waters and the risk that exposure to antibiotic resistant bacteria poses to human health. Curr. Opin. Microbiol. 2022, 65, 40–46.
- Stanton, I.C.; Bethel, A.; Leonard, A.F.C.; Gaze, W.H.; Garside, R. Existing evidence on antibiotic resistance exposure and transmission to humans from the environment: A systematic map. Environ. Evid. 2022, 11, 1–24.
- Zhang, X.-X.; Zhang, T.; Fang, H.H.P. Antibiotic resistance genes in water environment. Appl. Microbiol. Biotechnol. 2009, 82, 397–414.
- Rizzo, L.; Manaia, C.; Merlin, C.; Schwartz, T.; Dagot, C.; Ploy, M.C.; Michael, I.; Fatta-Kassinos, D. Urban wastewater treatment plants as hotspots for antibiotic resistant bacteria and genes spread into the environment: A review. Sci. Total Environ. 2013, 447, 345–360.
- Hendriksen, R.S.; Munk, P.; Njage, P.; van Bunnik, B.; McNally, L.; Lukjancenko, O.; Röder, T.; Nieuwenhuijse, D.; Pedersen, S.K.; Kjeldgaard, J.; et al. Global monitoring of antimicrobial resistance based on metagenomics analyses of urban sewage. Nat. Commun. 2019, 10, 1124.
- Pärnänen, K.M.M.; Narciso-da-Rocha, C.; Kneis, D.; Berendonk, T.U.; Cacace, D.; Do, T.T.; Elpers, C.; Fatta-Kassinos, D.; Henriques, I.; Jaeger, T.; et al. Resistance in European wastewater treatment plants mirrors the pattern of clinical antibiotic resistance prevalence. Sci. Adv. 2019, 5, eaau9124.
- Swami, O.C. Strategies to Combat Antimicrobial Resistance. J. Clin. Diagn. Res. 2014, 8, 8–11.
- WHO. Global Action Plan on Antimicrobial Resistance 2015. Available online: https://apps.who.int/iris/handle/10665/193736 (accessed on 10 May 2022).
- Berendonk, T.U.; Manaia, C.M.; Merlin, C.; Fatta-Kassinos, D.; Cytryn, E.; Walsh, F.; Bürgmann, H.; Sørum, H.; Norström, M.; Pons, M.-N.; et al. Tackling antibiotic resistance: The environmental framework. Nat. Rev. Microbiol. 2015, 13, 310–317.
- Barancheshme, F.; Munir, M. Strategies to Combat Antibiotic Resistance in the Wastewater Treatment Plants. Front. Microbiol. 2018, 8, 2603.
- Robinson, T.P.; Bu, D.P.; Carrique-Mas, J.; Fèvre, E.M.; Gilbert, M.; Grace, D.; Hay, S.I.; Jiwakanon, J.; Kakkar, M.; Kariuki, S.; et al. Antibiotic resistance is the quintessential One Health issue. Trans. R. Soc. Trop. Med. Hyg. 2016, 110, 377–380.
- Marano, R.B.M.; Fernandes, T.; Manaia, C.M.; Nunes, O.; Morrison, D.; Berendonk, T.U.; Kreuzinger, N.; Tenson, T.; Corno, G.; Fatta-Kassinos, D.; et al. A global multinational survey of cefotaxime-resistant coliforms in urban wastewater treatment plants. Environ. Int. 2020, 144, 106035.
- Larsson, D.G.J.; Flach, C.-F. Antibiotic resistance in the environment. Nat. Rev. Microbiol. 2022, 20, 257–269.
- Daoud, Z.; Rolain, J.-M. Editorial: “One Health” Approach for Revealing Reservoirs and Transmission of Antimicrobial Resistance. Front. Microbiol. 2022, 12, 2021–2023.
- Zhuang, M.; Achmon, Y.; Cao, Y.; Liang, X.; Chen, L.; Wang, H.; Siame, B.A.; Leung, K.Y. Distribution of antibiotic resistance genes in the environment. Environ. Pollut. 2021, 285, 117402.
- Sawa, T.; Kooguchi, K.; Moriyama, K. Molecular diversity of extended-spectrum β-lactamases and carbapenemases, and antimicrobial resistance. J. Intensive Care 2020, 8, 13.
- Baraniak, A.; Fiett, J.; Mrówka, A.; Walory, J.; Hryniewicz, W.; Gniadkowski, M. Evolution of TEM-Type Extended-Spectrum β-Lactamases in Clinical Enterobacteriaceae Strains in Poland. Antimicrob. Agents Chemother. 2005, 49, 1872–1880.
- De Champs, C.; Sirot, D.; Chanal, C.; Bonnet, R.; Sirot, J. A 1998 Survey of Extended-Spectrum β-Lactamases in Enterobacteriaceae in France. Antimicrob. Agents Chemother. 2000, 44, 3177–3179.
- Livermore, D.M.; Canton, R.; Gniadkowski, M.; Nordmann, P.; Rossolini, G.M.; Arlet, G.; Ayala, J.; Coque, T.M.; Kern-Zdanowicz, I.; Luzzaro, F.; et al. CTX-M: Changing the face of ESBLs in Europe. J. Antimicrob. Chemother. 2006, 59, 165–174.
- Doi, Y.; Iovleva, A.; Bonomo, R.A. The ecology of extended-spectrum β-lactamases (ESBLs) in the developed world. J. Travel Med. 2017, 24, S44–S51.
- Wiener, J. Multiple Antibiotic–Resistant Klebsiella and Escherichia coli in Nursing Homes. JAMA 1999, 281, 517.
- Pitout, J.D.D.; Nordmann, P.; Laupland, K.B.; Poirel, L. Emergence of Enterobacteriaceae producing extended-spectrum β-lactamases (ESBLs) in the community. J. Antimicrob. Chemother. 2005, 56, 52–59.
- Paterson, D.L.; Bonomo, R.A. Extended-Spectrum β-Lactamases: A Clinical Update. Clin. Microbiol. Rev. 2005, 18, 657–686.
- Nordmann, P.; Cuzon, G.; Naas, T. The real threat of Klebsiella pneumoniae carbapenemase-producing bacteria. Lancet Infect. Dis. 2009, 9, 228–236.
- Nordmann, P.; Naas, T.; Poirel, L. Global Spread of Carbapenemase-producing Enterobacteriaceae. Emerg. Infect. Dis. 2011, 17, 1791–1798.
- Munoz-Price, L.S.; Poirel, L.; Bonomo, R.A.; Schwaber, M.J.; Daikos, G.L.; Cormican, M.; Cornaglia, G.; Garau, J.; Gniadkowski, M.; Hayden, M.K.; et al. Clinical epidemiology of the global expansion of Klebsiella pneumoniae carbapenemases. Lancet Infect. Dis. 2013, 13, 785–796.
- Li, L.; Nesme, J.; Quintela-Baluja, M.; Balboa, S.; Hashsham, S.; Williams, M.R.; Yu, Z.; Sørensen, S.J.; Graham, D.W.; Romalde, J.L.; et al. Extended-Spectrum β-Lactamase and Carbapenemase Genes are Substantially and Sequentially Reduced during Conveyance and Treatment of Urban Sewage. Environ. Sci. Technol. 2021, 55, 5939–5949.
- Zaatout, N.; Bouras, S.; Slimani, N. Prevalence of extended-spectrum β-lactamase (ESBL)-producing Enterobacteriaceae in wastewater: A systematic review and meta-analysis. J. Water Health 2021, 19, 705–723.
- Gündoğdu, A.; Jennison, A.V.; Smith, H.V.; Stratton, H.; Katouli, M. Extended-spectrum β-lactamase producing Escherichia coli in hospital wastewaters and sewage treatment plants in Queensland, Australia. Can. J. Microbiol. 2013, 59, 737–745.
- Korzeniewska, E.; Harnisz, M. Extended-spectrum beta-lactamase (ESBL)-positive Enterobacteriaceae in municipal sewage and their emission to the environment. J. Environ. Manag. 2013, 128, 904–911.
- Makowska, N.; Philips, A.; Dabert, M.; Nowis, K.; Trzebny, A.; Koczura, R.; Mokracka, J. Metagenomic analysis of β-lactamase and carbapenemase genes in the wastewater resistome. Water Res. 2020, 170, 115277.
- Yuan, W.; Tian, T.; Yang, Q.; Riaz, L. Transfer potentials of antibiotic resistance genes in Escherichia spp. strains from different sources. Chemosphere 2020, 246, 125736.
- Ali, A.; Sultan, I.; Mondal, A.H.; Siddiqui, M.T.; Gogry, F.A.; Haq, Q.M.R. Lentic and effluent water of Delhi-NCR: A reservoir of multidrug-resistant bacteria harbouring bla CTX-M, bla TEM and bla SHV type ESBL genes. J. Water Health 2021, 19, 592–603.
- Fadare, F.T.; Okoh, A.I. The Abundance of Genes Encoding ESBL, pAmpC and Non-β-Lactam Resistance in Multidrug-Resistant Enterobacteriaceae Recovered from Wastewater Effluents. Front. Environ. Sci. 2021, 9, 1–12.
- Gumede, S.N.; Abia, A.L.K.; Amoako, D.G.; Essack, S.Y. Analysis of Wastewater Reveals the Spread of Diverse Extended-Spectrum β-Lactamase-Producing E. coli Strains in uMgungundlovu District, South Africa. Antibiotics 2021, 10, 860.
- Alouache, S.; Estepa, V.; Messai, Y.; Ruiz, E.; Torres, C.; Bakour, R. Characterization of ESBLs and Associated Quinolone Resistance in Escherichia coli and Klebsiella pneumoniae Isolates from an Urban Wastewater Treatment Plant in Algeria. Microb. Drug Resist. 2014, 20, 30–38.
- Bréchet, C.; Plantin, J.; Sauget, M.; Thouverez, M.; Talon, D.; Cholley, P.; Guyeux, C.; Hocquet, D.; Bertrand, X. Wastewater treatment plants release large amounts of extended-spectrum β-lactamase-producing escherichia coli into the environment. Clin. Infect. Dis. 2014, 58, 1658–1665.
- Ojer-Usoz, E.; González, D.; García-Jalón, I.; Vitas, A.I. High dissemination of extended-spectrum β-lactamase-producing Enterobacteriaceae in effluents from wastewater treatment plants. Water Res. 2014, 56, 37–47.
- Akiba, M.; Sekizuka, T.; Yamashita, A.; Kuroda, M.; Fujii, Y.; Murata, M.; Lee, K.I.; Joshua, D.I.; Balakrishna, K.; Bairy, I.; et al. Distribution and relationships of antimicrobial resistance determinants among extended-spectrum-cephalosporin-resistant or carbapenem-resistant Escherichia coli isolates from rivers and sewage treatment plants in India. Antimicrob. Agents Chemother. 2016, 60, 2972–2980.
- Caltagirone, M.; Nucleo, E.; Spalla, M.; Zara, F.; Novazzi, F.; Marchetti, V.M.; Piazza, A.; Bitar, I.; De Cicco, M.; Paolucci, S.; et al. Occurrence of Extended Spectrum β-Lactamases, KPC-Type, and MCR-1.2-Producing Enterobacteriaceae from Wells, River Water, and Wastewater Treatment Plants in Oltrepò Pavese Area, Northern Italy. Front. Microbiol. 2017, 8, 1–12.
- Conte, D.; Palmeiro, J.K.; da Silva Nogueira, K.; de Lima, T.M.R.; Cardoso, M.A.; Pontarolo, R.; Degaut Pontes, F.L.; Dalla-Costa, L.M. Characterization of CTX-M enzymes, quinolone resistance determinants, and antimicrobial residues from hospital sewage, wastewater treatment plant, and river water. Ecotoxicol. Environ. Saf. 2017, 136, 62–69.
- Guyomard-Rabenirina, S.; Dartron, C.; Falord, M.; Sadikalay, S.; Ducat, C.; Richard, V.; Breurec, S.; Gros, O.; Talarmin, A. Resistance to antimicrobial drugs in different surface waters and wastewaters of Guadeloupe. PLoS ONE 2017, 12, e0173155.
- Galler, H.; Feierl, G.; Petternel, C.; Reinthaler, F.; Haas, D.; Habib, J.; Kittinger, C.; Luxner, J.; Zarfel, G. Multiresistant Multiresistant Bacteria Isolated from Activated Sludge in Austria. Int. J. Environ. Res. Public Health 2018, 15, 479.
- Yang, Y.; Zhang, T.; Zhang, X.-X.; Liang, D.-W.; Zhang, M.; Gao, D.-W.; Zhu, H.-G.; Huang, Q.-G.; Fang, H.H.P. Quantification and characterization of β-lactam resistance genes in 15 sewage treatment plants from East Asia and North America. Appl. Microbiol. Biotechnol. 2012, 95, 1351–1358.
- Khan, M.A.; Thurgood, N.E.; Faheem, S.M.; Rais, N.; Ansari, M.Z.; Kaleem, S.M.; Khan, S.T. Occurrence of Extended Spectrum Beta-Lactamase Gram-Negative Bacteria from Non-Clinical Sources in Dubai, United Arab Emirates. Water 2020, 12, 2562.
- Osińska, A.; Korzeniewska, E.; Harnisz, M.; Felis, E.; Bajkacz, S.; Jachimowicz, P.; Niestępski, S.; Konopka, I. Small-scale wastewater treatment plants as a source of the dissemination of antibiotic resistance genes in the aquatic environment. J. Hazard. Mater. 2020, 381, 121221.
- Surleac, M.; Barbu, I.C.; Paraschiv, S.; Popa, L.I.; Gheorghe, I.; Marutescu, L.; Popa, M.; Sarbu, I.; Talapan, D.; Nita, M.; et al. Whole genome sequencing snapshot of multidrug resistant Klebsiella pneumoniae strains from hospitals and receiving wastewater treatment plants in Southern Romania. PLoS ONE 2020, 15, e0228079.
- Smyth, C.; O’Flaherty, A.; Walsh, F.; Do, T.T. Antibiotic resistant and extended-spectrum β-lactamase producing faecal coliforms in wastewater treatment plant effluent. Environ. Pollut. 2020, 262, 114244.
- Aristizábal-Hoyos, A.M.; Rodríguez, E.A.; Arias, L.; Jiménez, J.N. High clonal diversity of multidrug-resistant and extended spectrum beta-lactamase-producing Escherichia coli in a wastewater treatment plant. J. Environ. Manag. 2019, 245, 37–47.
- Amador, P.P.; Fernandes, R.M.; Prudêncio, M.C.; Barreto, M.P.; Duarte, I.M. Antibiotic resistance in wastewater: Occurrence and fate of Enterobacteriaceae producers of Class A and Class C β-lactamases. J. Environ. Sci. Health Part A 2015, 50, 26–39.
- Proia, L.; Anzil, A.; Borrego, C.; Farrè, M.; Llorca, M.; Sanchis, J.; Bogaerts, P.; Balcázar, J.L.; Servais, P. Occurrence and persistence of carbapenemases genes in hospital and wastewater treatment plants and propagation in the receiving river. J. Hazard. Mater. 2018, 358, 33–43.
- Zhang, A.; Call, D.R.; Besser, T.E.; Liu, J.; Jones, L.; Wang, H.; Davis, M.A. β-lactam resistance genes in bacteriophage and bacterial DNA from wastewater, river water, and irrigation water in Washington State. Water Res. 2019, 161, 335–340.
- Adegoke, A.A.; Madu, C.E.; Aiyegoro, O.A.; Stenström, T.A.; Okoh, A.I. Antibiogram and beta-lactamase genes among cefotaxime resistant E. coli from wastewater treatment plant. Antimicrob. Resist. Infect. Control 2020, 9, 46.
- Piotrowska, M.; Kowalska, S.; Popowska, M. Diversity of β-lactam resistance genes in gram-negative rods isolated from a municipal wastewater treatment plant. Ann. Microbiol. 2019, 69, 591–601.
- Hubeny, J.; Ciesielski, S.; Harnisz, M.; Korzeniewska, E.; Dulski, T.; Jałowiecki, Ł.; Płaza, G. Impact of Hospital Wastewater on the Occurrence and Diversity of Beta-Lactamase Genes During Wastewater Treatment with an Emphasis on Carbapenemase Genes: A Metagenomic Approach. Front. Environ. Sci. 2021, 9, 738158.
- Teban-Man, A.; Farkas, A.; Baricz, A.; Hegedus, A.; Szekeres, E.; Pârvu, M.; Coman, C. Wastewaters, with or without Hospital Contribution, Harbour MDR, Carbapenemase-Producing, but Not Hypervirulent Klebsiella pneumoniae. Antibiotics 2021, 10, 361.
- Galler, H.; Feierl, G.; Petternel, C.; Reinthaler, F.F.; Haas, D.; Grisold, A.J.; Luxner, J.; Zarfel, G. KPC-2 and OXA-48 carbapenemase-harbouring Enterobacteriaceae detected in an Austrian wastewater treatment plant. Clin. Microbiol. Infect. 2014, 20, O132–O134.
- Bengtsson-Palme, J.; Hammarén, R.; Pal, C.; Östman, M.; Björlenius, B.; Flach, C.-F.; Fick, J.; Kristiansson, E.; Tysklind, M.; Larsson, D.G.J. Elucidating selection processes for antibiotic resistance in sewage treatment plants using metagenomics. Sci. Total Environ. 2016, 572, 697–712.
- Piotrowska, M.; Przygodzińska, D.; Matyjewicz, K.; Popowska, M. Occurrence and Variety of β-Lactamase Genes among Aeromonas spp. Isolated from Urban Wastewater Treatment Plant. Front. Microbiol. 2017, 8, 863.
- Subirats, J.; Royo, E.; Balcázar, J.L.; Borrego, C.M. Real-time PCR assays for the detection and quantification of carbapenemase genes (bla KPC, bla NDM, and bla OXA-48) in environmental samples. Environ. Sci. Pollut. Res. 2017, 24, 6710–6714.
- Zurfluh, K.; Bagutti, C.; Brodmann, P.; Alt, M.; Schulze, J.; Fanning, S.; Stephan, R.; Nüesch-Inderbinen, M. Wastewater is a reservoir for clinically relevant carbapenemase- and 16s rRNA methylase-producing Enterobacteriaceae. Int. J. Antimicrob. Agents 2017, 50, 436–440.
- Müller, H.; Sib, E.; Gajdiss, M.; Klanke, U.; Lenz-Plet, F.; Barabasch, V.; Albert, C.; Schallenberg, A.; Timm, C.; Zacharias, N.; et al. Dissemination of multi-resistant Gram-negative bacteria into German wastewater and surface waters. FEMS Microbiol. Ecol. 2018, 94, fiy057.
- Cacace, D.; Fatta-Kassinos, D.; Manaia, C.M.; Cytryn, E.; Kreuzinger, N.; Rizzo, L.; Karaolia, P.; Schwartz, T.; Alexander, J.; Merlin, C.; et al. Antibiotic resistance genes in treated wastewater and in the receiving water bodies: A pan-European survey of urban settings. Water Res. 2019, 162, 320–330.
- Mathys, D.A.; Mollenkopf, D.F.; Feicht, S.M.; Adams, R.J.; Albers, A.L.; Stuever, D.M.; Grooters, S.V.; Ballash, G.A.; Daniels, J.B.; Wittum, T.E. Carbapenemase-producing Enterobacteriaceae and Aeromonas spp. Present in wastewater treatment plant effluent and nearby surface waters in the US. PLoS ONE 2018, 14, e0218650.
- Hoelle, J.; Johnson, J.R.; Johnston, B.D.; Kinkle, B.; Boczek, L.; Ryu, H.; Hayes, S. Survey of US wastewater for carbapenem-resistant Enterobacteriaceae. J. Water Health 2019, 17, 219–226.
- Cooper, A.L.; Carter, C.; McLeod, H.; Wright, M.; Sritharan, P.; Tamber, S.; Wong, A.; Carrillo, C.D.; Blais, B.W. Detection of carbapenem-resistance genes in bacteria isolated from wastewater in Ontario. FACETS 2021, 6, 569–591.
- Ebomah, K.E.; Okoh, A.I. Detection of Carbapenem-Resistance Genes in Klebsiella Species Recovered from Selected Environmental Niches in the Eastern Cape Province, South Africa. Antibiotics 2020, 9, 425.
- Yang, F.; Mao, D.; Zhou, H.; Luo, Y. Prevalence and Fate of Carbapenemase Genes in a Wastewater Treatment Plant in Northern China. PLoS ONE 2016, 11, e0156383.
- Ng, C.; Tan, B.; Jiang, X.-T.; Gu, X.; Chen, H.; Schmitz, B.W.; Haller, L.; Charles, F.R.; Zhang, T.; Gin, K. Metagenomic and Resistome Analysis of a Full-Scale Municipal Wastewater Treatment Plant in Singapore Containing Membrane Bioreactors. Front. Microbiol. 2019, 10, 172.
- Ng, C.; Tay, M.; Tan, B.; Le, T.-H.; Haller, L.; Chen, H.; Koh, T.H.; Barkham, T.M.S.; Thompson, J.R.; Gin, K.Y.H. Characterization of Metagenomes in Urban Aquatic Compartments Reveals High Prevalence of Clinically Relevant Antibiotic Resistance Genes in Wastewaters. Front. Microbiol. 2017, 8, 2200.
- Khan, F.A.; Söderquist, B.; Jass, J. Prevalence and Diversity of Antibiotic Resistance Genes in Swedish Aquatic Environments Impacted by Household and Hospital Wastewater. Front. Microbiol. 2019, 10, 688.
- Araújo, S.; Sousa, M.; Tacão, M.; Baraúna, R.A.; Silva, A.; Ramos, R.; Alves, A.; Manaia, C.M.; Henriques, I. Carbapenem-resistant bacteria over a wastewater treatment process: Carbapenem-resistant Enterobacteriaceae in untreated wastewater and intrinsically-resistant bacteria in final effluent. Sci. Total Environ. 2021, 782, 146892.
- Zhang, D.; Peng, Y.; Chan, C.-L.; On, H.; Wai, H.K.-F.; Shekhawat, S.S.; Gupta, A.B.; Varshney, A.K.; Chuanchuen, R.; Zhou, X.; et al. Metagenomic Survey Reveals More Diverse and Abundant Antibiotic Resistance Genes in Municipal Wastewater Than Hospital Wastewater. Front. Microbiol. 2021, 12, 712843.
- Reinthaler, F.F.; Feierl, G.; Galler, H.; Haas, D.; Leitner, E.; Mascher, F.; Melkes, A.; Posch, J.; Winter, I.; Zarfel, G. ESBL-producing E. coli in Austrian sewage sludge. Water Res. 2010, 44, 1981–1985.
- Dolejska, M.; Frolkova, P.; Florek, M.; Jamborova, I.; Purgertova, M.; Kutilova, I.; Cizek, A.; Guenther, S.; Literak, I. CTX-M-15-producing Escherichia coli clone B2-O25b-ST131 and Klebsiella spp. isolates in municipal wastewater treatment plant effluents. J. Antimicrob. Chemother. 2011, 66, 2784–2790.
- Korzeniewska, E.; Korzeniewska, A.; Harnisz, M. Antibiotic resistant Escherichia coli in hospital and municipal sewage and their emission to the environment. Ecotoxicol. Environ. Saf. 2013, 91, 96–102.
- Čornejová, T.; Venglovsky, J.; Gregova, G.; Kmetova, M.; Kmet, V. Extended spectrum beta-lactamases in Escherichia coli from municipal wastewater. Ann. Agric. Environ. Med. 2015, 22, 447–450.
- Kwak, Y.-K.; Colque, P.; Byfors, S.; Giske, C.G.; Möllby, R.; Kühn, I. Surveillance of antimicrobial resistance among Escherichia coli in wastewater in Stockholm during 1 year: Does it reflect the resistance trends in the society? Int. J. Antimicrob. Agents 2015, 45, 25–32.
- Dropa, M.; Lincopan, N.; Balsalobre, L.C.; Oliveira, D.E.; Moura, R.A.; Fernandes, M.R.; da Silva, Q.M.; Matté, G.R.; Sato, M.I.Z.; Matté, M.H. Genetic background of novel sequence types of CTX-M-8- and CTX-M-15-producing Escherichia coli and Klebsiella pneumoniae from public wastewater treatment plants in São Paulo, Brazil. Environ. Sci. Pollut. Res. 2016, 23, 4953–4958.
- Silva, I.; Tacão, M.; Tavares, R.D.S.; Miranda, R.; Araújo, S.; Manaia, C.M.; Henriques, I. Fate of cefotaxime-resistant Enterobacteriaceae and ESBL-producers over a full-scale wastewater treatment process with UV disinfection. Sci. Total Environ. 2018, 639, 1028–1037.
- Haberecht, H.B.; Nealon, N.J.; Gilliland, J.R.; Holder, A.V.; Runyan, C.; Oppel, R.C.; Ibrahim, H.M.; Mueller, L.; Schrupp, F.; Vilchez, S.; et al. Antimicrobial-Resistant Escherichia coli from Environmental Waters in Northern Colorado. J. Environ. Public Health 2019, 2019, 1–13.
- Raven, K.E.; Ludden, C.; Gouliouris, T.; Blane, B.; Naydenova, P.; Brown, N.M.; Parkhill, J.; Peacock, S.J. Genomic surveillance of Escherichia coli in municipal wastewater treatment plants as an indicator of clinically relevant pathogens and their resistance genes. Microb. Genom. 2019, 5, e000267.
- Sghaier, S.; Abbassi, M.S.; Pascual, A.; Serrano, L.; Díaz-De-Alba, P.; Said, M.B.; Hassen, B.; Ibrahim, C.; Hassen, A.; López-Cerero, L. Extended-spectrum β-lactamase-producing Enterobacteriaceae from animal origin and wastewater in Tunisia: First detection of O25b-B23-CTX-M-27-ST131 Escherichia coli and CTX-M-15/OXA-204-producing Citrobacter freundii from wastewater. J. Glob. Antimicrob. Resist. 2019, 17, 189–194.
- Tanaka, H.; Hayashi, W.; Iimura, M.; Taniguchi, Y.; Soga, E.; Matsuo, N.; Kawamura, K.; Arakawa, Y.; Nagano, Y.; Nagano, N. Wastewater as a Probable Environmental Reservoir of Extended-Spectrum-β-Lactamase Genes: Detection of Chimeric β-Lactamases CTX-M-64 and CTX-M-123. Appl. Environ. Microbiol. 2019, 85, 1–12.
- Schages, L.; Wichern, F.; Kalscheuer, R.; Bockmühl, D. Winter is coming—Impact of temperature on the variation of beta-lactamase and mcr genes in a wastewater treatment plant. Sci. Total Environ. 2020, 712, 136499.
- Urano, N.; Okai, M.; Tashiro, Y.; Takeuchi, A.; Endo, R.; Ishida, M.; Takashio, M. Behavior of Antibiotic-Resistant Fecal Coliforms in the Stream of a Sewage Treatment Plant in Tokyo. Adv. Microbiol. 2020, 10, 318–330.
- Liedhegner, E.; Bojar, B.; Beattie, R.E.; Cahak, C.; Hristova, K.R.; Skwor, T. Similarities in Virulence and Extended Spectrum Beta-Lactamase Gene Profiles among Cefotaxime-Resistant Escherichia coli Wastewater and Clinical Isolates. Antibiotics 2022, 11, 260.
- Igbinosa, I.H.; Okoh, A.I. Antibiotic Susceptibility Profile of Aeromonas Species Isolated from Wastewater Treatment Plant. Sci. World J. 2012, 2012, 1–6.
- Ben Said, L.; Jouini, A.; Alonso, C.A.; Klibi, N.; Dziri, R.; Boudabous, A.; Ben Slama, K.; Torres, C. Characteristics of extended-spectrum β-lactamase (ESBL)- and pAmpC beta-lactamase-producing Enterobacteriaceae of water samples in Tunisia. Sci. Total Environ. 2016, 550, 1103–1109.
- Ludden, C.; Reuter, S.; Judge, K.; Gouliouris, T.; Blane, B.; Coll, F.; Naydenova, P.; Hunt, M.; Tracey, A.; Hopkins, K.L.; et al. Sharing of carbapenemase-encoding plasmids between Enterobacteriaceae in UK sewage uncovered by MinION sequencing. Microb. Genom. 2017, 3, e000114.
- Zhang, S.; Han, B.; Gu, J.; Wang, C.; Wang, P.; Ma, Y.; Cao, J.; He, Z. Fate of antibiotic resistant cultivable heterotrophic bacteria and antibiotic resistance genes in wastewater treatment processes. Chemosphere 2015, 135, 138–145.
- Nzima, B.; Adegoke, A.A.; Ofon, U.A.; Al-Dahmoshi, H.O.M.; Saki, M.; Ndubuisi-Nnaji, U.U.; Inyang, C.U. Resistotyping and extended-spectrum beta-lactamase genes among Escherichia coli from wastewater treatment plants and recipient surface water for reuse in South Africa. New Microbes New Infect. 2020, 38, 100803.
- Gomi, R.; Matsuda, T.; Matsumura, Y.; Yamamoto, M.; Tanaka, M.; Ichiyama, S.; Yoneda, M. Occurrence of clinically important lineages, including the sequence type 131 C1-M27 subclone, among extended-spectrum--lactamase-producing Escherichia coli in wastewater. Antimicrob. Agents Chemother. 2017, 61, e00564-17.
- Finn, T.J.; Scriver, L.; Lam, L.; Duong, M.; Peirano, G.; Lynch, T.; Dong, T.; Pitout, J.D.D.; DeVinney, R. A comprehensive account of escherichia coli sequence type 131 in wastewater reveals an abundance of fluoroquinolone-resistant clade a strains. Appl. Environ. Microbiol. 2020, 86, 1–11.
- Zhi, S.; Stothard, P.; Banting, G.; Scott, C.; Huntley, K.; Ryu, K.; Otto, S.; Ashbolt, N.; Checkley, S.; Dong, T.; et al. Characterization of water treatment-resistant and multidrug-resistant urinary pathogenic Escherichia coli in treated wastewater. Water Res. 2020, 182, 115827.
- Mesquita, E.; Ribeiro, R.; Silva, C.J.C.; Alves, R.; Baptista, R.; Condinho, S.; Rosa, M.J.; Perdigão, J.; Caneiras, C.; Duarte, A. An update on wastewater multi-resistant bacteria: Identification of clinical pathogens such as escherichia coli o25b:H4-b2-st131-producing ctx-m-15 esbl and kpc-3 carbapenemase-producing klebsiella oxytoca. Microorganisms 2021, 9, 576.
- Hembach, N.; Schmid, F.; Alexander, J.; Hiller, C.; Rogall, E.T.; Schwartz, T. Occurrence of the mcr-1 Colistin Resistance Gene and other Clinically Relevant Antibiotic Resistance Genes in Microbial Populations at Different Municipal Wastewater Treatment Plants in Germany. Front. Microbiol. 2017, 8, 1282.
- Szczepanowski, R.; Linke, B.; Krahn, I.; Gartemann, K.-H.; Gützkow, T.; Eichler, W.; Pühler, A.; Schlüter, A. Detection of 140 clinically relevant antibiotic-resistance genes in the plasmid metagenome of wastewater treatment plant bacteria showing reduced susceptibility to selected antibiotics. Microbiology 2009, 155, 2306–2319.
- Wen, Q.; Yang, L.; Duan, R.; Chen, Z. Monitoring and evaluation of antibiotic resistance genes in four municipal wastewater treatment plants in Harbin, Northeast China. Environ. Pollut. 2016, 212, 34–40.
- Narciso-da-Rocha, C.; Rocha, J.; Vaz-Moreira, I.; Lira, F.; Tamames, J.; Henriques, I.; Martinez, J.L.; Manaia, C.M. Bacterial lineages putatively associated with the dissemination of antibiotic resistance genes in a full-scale urban wastewater treatment plant. Environ. Int. 2018, 118, 179–188.
- Neudorf, K.D.; Huang, Y.N.; Ragush, C.M.; Yost, C.K.; Jamieson, R.C.; Truelstrup Hansen, L. Antibiotic resistance genes in municipal wastewater treatment systems and receiving waters in Arctic Canada. Sci. Total Environ. 2017, 598, 1085–1094.
- Rodriguez-Mozaz, S.; Chamorro, S.; Marti, E.; Huerta, B.; Gros, M.; Sànchez-Melsió, A.; Borrego, C.M.; Barceló, D.; Balcázar, J.L. Occurrence of antibiotics and antibiotic resistance genes in hospital and urban wastewaters and their impact on the receiving river. Water Res. 2015, 69, 234–242.
- Laht, M.; Karkman, A.; Voolaid, V.; Ritz, C.; Tenson, T.; Virta, M.; Kisand, V. Abundances of Tetracycline, Sulphonamide and Beta-Lactam Antibiotic Resistance Genes in Conventional Wastewater Treatment Plants (WWTPs) with Different Waste Load. PLoS ONE 2014, 9, e103705.
- Rafraf, I.D.; Lekunberri, I.; Sànchez-Melsió, A.; Aouni, M.; Borrego, C.M.; Balcázar, J.L. Abundance of antibiotic resistance genes in five municipal wastewater treatment plants in the Monastir Governorate, Tunisia. Environ. Pollut. 2016, 219, 353–358.
- Kowalski, M.; Wolany, J.; Pastuszka, J.S.; Płaza, G.; Wlazło, A.; Ulfig, K.; Malina, A. Characteristics of airborne bacteria and fungi in some Polish wastewater treatment plants. Int. J. Environ. Sci. Technol. 2017, 14, 2181–2192.
- Ginn, O.; Rocha-Melogno, L.; Bivins, A.; Lowry, S.; Cardelino, M.; Nichols, D.; Tripathi, S.N.; Soria, F.; Andrade, M.; Bergin, M.; et al. Detection and Quantification of Enteric Pathogens in Aerosols near Open Wastewater Canals in Cities with Poor Sanitation. Environ. Sci. Technol. 2021, 55, 14758–14771.
- Wengenroth, L.; Berglund, F.; Blaak, H.; Chifiriuc, M.C.; Flach, C.-F.; Pircalabioru, G.G.; Larsson, D.G.J.; Marutescu, L.; van Passel, M.W.J.; Popa, M.; et al. Antibiotic Resistance in Wastewater Treatment Plants and Transmission Risks for Employees and Residents: The Concept of the AWARE Study. Antibiotics 2021, 10, 478.
- Lee, G.; Yoo, K. A review of the emergence of antibiotic resistance in bioaerosols and its monitoring methods. Rev. Environ. Sci. Bio/Technol. 2022, 21, 799–827.
- Gaviria-Figueroa, A.; Preisner, E.C.; Hoque, S.; Feigley, C.E.; Norman, R.S. Emission and dispersal of antibiotic resistance genes through bioaerosols generated during the treatment of municipal sewage. Sci. Total Environ. 2019, 686, 402–412.
- Palzkill, T. Metallo-β-lactamase structure and function. Ann. N. Y. Acad. Sci. 2013, 1277, 91–104.
- Khan, A.U.; Maryam, L.; Zarrilli, R. Structure, Genetics and Worldwide Spread of New Delhi Metallo-β-lactamase (NDM): A threat to public health. BMC Microbiol. 2017, 17, 101.
- Cui, X.; Zhang, H.; Du, H. Carbapenemases in Enterobacteriaceae: Detection and Antimicrobial Therapy. Front. Microbiol. 2019, 10, 1823.
- Kazmierczak, K.M.; Rabine, S.; Hackel, M.; McLaughlin, R.E.; Biedenbach, D.J.; Bouchillon, S.K.; Sahm, D.F.; Bradford, P.A. Multiyear, Multinational Survey of the Incidence and Global Distribution of Metallo-β-Lactamase-Producing Enterobacteriaceae and Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2016, 60, 1067–1078.
- Matsumura, Y.; Peirano, G.; Motyl, M.R.; Adams, M.D.; Chen, L.; Kreiswirth, B.; DeVinney, R.; Pitout, J.D.D. Global Molecular Epidemiology of IMP-Producing Enterobacteriaceae. Antimicrob. Agents Chemother. 2017, 61, e02729-16.
- Matsumura, Y.; Peirano, G.; Bradford, P.A.; Motyl, M.R.; DeVinney, R.; Pitout, J.D.D. Genomic characterization of IMP and VIM carbapenemase-encoding transferable plasmids of Enterobacteriaceae. J. Antimicrob. Chemother. 2018, 73, 3034–3038.
- Ghaith, D.M.; Zafer, M.M.; Ismail, D.K.; Al-Agamy, M.H.; Bohol, M.F.F.; Al-Qahtani, A.; Al-Ahdal, M.N.; Elnagdy, S.M.; Mostafa, I.Y. First reported nosocomial outbreak of Serratia marcescens harboring blaIMP-4 and blaVIM-2 in a neonatal intensive care unit in Cairo, Egypt. Infect. Drug Resist. 2018, 11, 2211–2217.
- Castanheira, M.; Deshpande, L.M.; Mendes, R.E.; Canton, R.; Sader, H.S.; Jones, R.N. Variations in the Occurrence of Resistance Phenotypes and Carbapenemase Genes Among Enterobacteriaceae Isolates in 20 Years of the SENTRY Antimicrobial Surveillance Program. Open Forum Infect. Dis. 2019, 6, S23–S33.
- Pournaras, S. Hospital outbreak of multiple clones of Pseudomonas aeruginosa carrying the unrelated metallo-beta-lactamase gene variants blaVIM-2 and blaVIM-4. J. Antimicrob. Chemother. 2003, 51, 1409–1414.
- Lagatolla, C.; Edalucci, E.; Dolzani, L.; Riccio, M.L.; De Luca, F.; Medessi, E.; Rossolini, G.M.; Tonin, E.A. Molecular Evolution of Metallo-β-Lactamase-Producing Pseudomonas aeruginosa in a Nosocomial Setting of High-Level Endemicity. J. Clin. Microbiol. 2006, 44, 2348–2353.
- Yong, D.; Toleman, M.A.; Giske, C.G.; Cho, H.S.; Sundman, K.; Lee, K.; Walsh, T.R. Characterization of a new metallo-β-lactamase gene, bla NDM-1, and a novel erythromycin esterase gene carried on a unique genetic structure in Klebsiella pneumoniae sequence type 14 from India. Antimicrob. Agents Chemother. 2009, 53, 5046–5054.
- Bush, K.; Bradford, P.A. Epidemiology of β-Lactamase-Producing Pathogens. Clin. Microbiol. Rev. 2020, 33, 1–37.
- van Loon, K.; Voor in ‘t holt, A.F.; Vos, M.C. A Systematic Review and Meta-analyses of the Clinical Epidemiology of Carbapenem-Resistant Enterobacteriaceae. Antimicrob. Agents Chemother. 2018, 62, 1–18.
- Wu, W.; Feng, Y.; Tang, G.; Qiao, F.; McNally, A.; Zong, Z. NDM Metallo-β-Lactamases and Their Bacterial Producers in Health Care Settings. Clin. Microbiol. Rev. 2019, 32, e00115-18.
- Hansen, G.T. Continuous Evolution: Perspective on the Epidemiology of Carbapenemase Resistance Among Enterobacterales and Other Gram-Negative Bacteria. Infect. Dis. Ther. 2021, 10, 75–92.
- Alexander, J.; Bollmann, A.; Seitz, W.; Schwartz, T. Microbiological characterization of aquatic microbiomes targeting taxonomical marker genes and antibiotic resistance genes of opportunistic bacteria. Sci. Total Environ. 2015, 512–513, 316–325.
- Hubeny, J.; Korzeniewska, E.; Buta-Hubeny, M.; Zieliński, W.; Rolbiecki, D.; Harnisz, M. Characterization of carbapenem resistance in environmental samples and Acinetobacter spp. isolates from wastewater and river water in Poland. Sci. Total Environ. 2022, 822, 153437.
- Khan, F.A.; Hellmark, B.; Ehricht, R.; Söderquist, B.; Jass, J. Related carbapenemase-producing Klebsiella isolates detected in both a hospital and associated aquatic environment in Sweden. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 2241–2251.
- Mills, M.C.; Lee, J. The threat of carbapenem-resistant bacteria in the environment: Evidence of widespread contamination of reservoirs at a global scale. Environ. Pollut. 2019, 255, 113143.
- Cherak, Z.; Loucif, L.; Moussi, A.; Rolain, J.-M. Carbapenemase-producing Gram-negative bacteria in aquatic environments: A review. J. Glob. Antimicrob. Resist. 2021, 25, 287–309.
- Ranjan, R.; Thatikonda, S. β-Lactam Resistance Gene NDM-1 in the Aquatic Environment: A Review. Curr. Microbiol. 2021, 78, 3634–3643.
- Lamba, M.; Graham, D.W.; Ahammad, S.Z. Hospital Wastewater Releases of Carbapenem-Resistance Pathogens and Genes in Urban India. Environ. Sci. Technol. 2017, 51, 13906–13912.
- Parvez, S.; Khan, A.U. Hospital sewage water: A reservoir for variants of New Delhi metallo-β-lactamase (NDM)- and extended-spectrum β-lactamase (ESBL)-producing Enterobacteriaceae. Int. J. Antimicrob. Agents 2018, 51, 82–88.
- Bardhan, T.; Chakraborty, M.; Bhattacharjee, B. Prevalence of colistin-resistant, carbapenem-hydrolyzing proteobacteria in hospital water bodies and out-falls of West Bengal, India. Int. J. Environ. Res. Public Health 2020, 17, 1007.
- Hwang, S.H.; Kim, Y.J. Meropenem-resistant bacteria in hospital effluents in Seoul, Korea. Environ. Monit. Assess. 2018, 190, 673.
- Haller, L.; Chen, H.; Ng, C.; Le, T.H.; Koh, T.H.; Barkham, T.; Sobsey, M.; Gin, K.Y.H. Occurrence and characteristics of extended-spectrum β-lactamase- and carbapenemase- producing bacteria from hospital effluents in Singapore. Sci. Total Environ. 2018, 615, 1119–1125.
- Suzuki, Y.; Nazareno, P.J.; Nakano, R.; Mondoy, M.; Nakano, A.; Bugayong, M.P.; Bilar, J.; Perez, M.; Medina, E.J.; Saito-Obata, M.; et al. Environmental Presence and Genetic Characteristics of Carbapenemase-Producing Enterobacteriaceae from Hospital Sewage and River Water in the Philippines. Appl. Environ. Microbiol. 2020, 86, 1–10.
- Nasri, E.; Subirats, J.; Sànchez-Melsió, A.; Ben Mansour, H.; Borrego, C.M.; Balcázar, J.L. Abundance of carbapenemase genes (blaKPC, blaNDM and blaOXA-48) in wastewater effluents from Tunisian hospitals. Environ. Pollut. 2017, 229, 371–374.
- Marathe, N.P.; Berglund, F.; Razavi, M.; Pal, C.; Dröge, J.; Samant, S.; Kristiansson, E.; Larsson, D.G.J. Sewage effluent from an Indian hospital harbors novel carbapenemases and integron-borne antibiotic resistance genes. Microbiome 2019, 7, 97.
- Zhang, L.; Ma, X.; Luo, L.; Hu, N.; Duan, J.; Tang, Z.; Zhong, R.; Li, Y. The Prevalence and Characterization of Extended-Spectrum β-Lactamase- and Carbapenemase-Producing Bacteria from Hospital Sewage, Treated Effluents and Receiving Rivers. Int. J. Environ. Res. Public Health 2020, 17, 1183.
- Luo, Y.; Yang, F.; Mathieu, J.; Mao, D.; Wang, Q.; Alvarez, P.J.J. Proliferation of Multidrug-Resistant New Delhi Metallo-β-lactamase Genes in Municipal Wastewater Treatment Plants in Northern China. Environ. Sci. Technol. Lett. 2014, 1, 26–30.
- Yang, F.; Mao, D.; Zhou, H.; Wang, X.; Luo, Y. Propagation of New Delhi Metallo-β-lactamase Genes (bla NDM-1) from a Wastewater Treatment Plant to Its Receiving River. Environ. Sci. Technol. Lett. 2016, 3, 138–143.
- Mantilla-Calderon, D.; Jumat, M.R.; Wang, T.; Ganesan, P.; Al-Jassim, N.; Hong, P.Y. Isolation and characterization of NDM-positive Escherichia coli from municipal wastewater in Jeddah, Saudi Arabia. Antimicrob. Agents Chemother. 2016, 60, 5223–5231.
- Stachurová, T.; Piková, H.; Bartas, M.; Semerád, J.; Svobodová, K.; Malachová, K. Beta-lactam resistance development during the treatment processes of municipal wastewater treatment plants. Chemosphere 2021, 280, 130749.
- Mahon, B.M.; Brehony, C.; McGrath, E.; Killeen, J.; Cormican, M.; Hickey, P.; Keane, S.; Hanahoe, B.; Dolan, A.; Morris, D. Indistinguishable NDM-producing Escherichia coli isolated from recreational waters, sewage, and a clinical specimen in Ireland, 2016 to 2017. Eurosurveillance 2017, 22, 30513.
- Walsh, T.R.; Weeks, J.; Livermore, D.M.; Toleman, M.A. Dissemination of NDM-1 positive bacteria in the New Delhi environment and its implications for human health: An environmental point prevalence study. Lancet Infect. Dis. 2011, 11, 355–362.
- Lamba, M.; Gupta, S.; Shukla, R.; Graham, D.W.; Sreekrishnan, T.R.; Ahammad, S.Z. Carbapenem resistance exposures via wastewaters across New Delhi. Environ. Int. 2018, 119, 302–308.
- Divyashree, M.; Mani, M.K.; Shama Prakash, K.; Vijaya Kumar, D.; Veena Shetty, A.; Shetty, A.K.; Karunasagar, I. Hospital wastewater treatment reduces NDM-positive bacteria being discharged into water bodies. Water Environ. Res. 2020, 92, 562–568.
- Bonomo, R.A. β-Lactamases: A Focus on Current Challenges. Cold Spring Harb. Perspect. Med. 2017, 7, a025239.
- Philippon, A.; Arlet, G.; Jacoby, G.A. Plasmid-Determined AmpC-Type β-Lactamases. Antimicrob. Agents Chemother. 2002, 46, 1–11. Bauernfeind, A.; Stemplinger, I.; Jungwirth, R.; Wilhelm, R.; Chong, Y. Comparative characterization of the cephamycinase blaCMY-1 gene and its relationship with other beta-lactamase genes. Antimicrob. Agents Chemother. 1996, 40, 1926–1930.
- Jacoby, G.A. AmpC β-Lactamases. Clin. Microbiol. Rev. 2009, 22, 161–182. Philippon, A.; Arlet, G.; Jacoby, G.A. Plasmid-Determined AmpC-Type β-Lactamases. Antimicrob. Agents Chemother. 2002, 46, 1–11.
- Pérez-Etayo, L.; González, D.; Leiva, J.; Vitas, A.I. Multidrug-Resistant Bacteria Isolated from Different Aquatic Environments in the North of Spain and South of France. Microorganisms 2020, 8, 1425. Jacoby, G.A. AmpC β-Lactamases. Clin. Microbiol. Rev. 2009, 22, 161–182.
- Yim, G.; Kwong, W.; Davies, J.; Miao, V. Complex integrons containing qnrB4-ampC (bla DHA-1) in plasmids of multidrug-resistant Citrobacter freundii from wastewater. Can. J. Microbiol. 2013, 59, 110–116. Pérez-Etayo, L.; González, D.; Leiva, J.; Vitas, A.I. Multidrug-Resistant Bacteria Isolated from Different Aquatic Environments in the North of Spain and South of France. Microorganisms 2020, 8, 1425.
- Su, H.-C.; Ying, G.-G.; He, L.-Y.; Liu, Y.-S.; Zhang, R.-Q.; Tao, R. Antibiotic resistance, plasmid-mediated quinolone resistance (PMQR) genes and ampC gene in two typical municipal wastewater treatment plants. Environ. Sci. Process. Impacts 2014, 16, 324. Yim, G.; Kwong, W.; Davies, J.; Miao, V. Complex integrons containing qnrB4-ampC (bla DHA-1) in plasmids of multidrug-resistant Citrobacter freundii from wastewater. Can. J. Microbiol. 2013, 59, 110–116.
- Pitout, J.D.D.; Peirano, G.; Kock, M.M.; Strydom, K.; Matsumura, Y. The Global Ascendency of OXA-48-Type Carbapenemases. Clin. Microbiol. Rev. 2019, 33, 1–48. Su, H.-C.; Ying, G.-G.; He, L.-Y.; Liu, Y.-S.; Zhang, R.-Q.; Tao, R. Antibiotic resistance, plasmid-mediated quinolone resistance (PMQR) genes and ampC gene in two typical municipal wastewater treatment plants. Environ. Sci. Process. Impacts 2014, 16, 324.
- Evans, B.A.; Amyes, S.G.B. OXA β-Lactamases. Clin. Microbiol. Rev. 2014, 27, 241–263. Pitout, J.D.D.; Peirano, G.; Kock, M.M.; Strydom, K.; Matsumura, Y. The Global Ascendency of OXA-48-Type Carbapenemases. Clin. Microbiol. Rev. 2019, 33, 1–48.
- Mairi, A.; Pantel, A.; Sotto, A.; Lavigne, J.-P.; Touati, A. OXA-48-like carbapenemases producing Enterobacteriaceae in different niches. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 587–604. Evans, B.A.; Amyes, S.G.B. OXA β-Lactamases. Clin. Microbiol. Rev. 2014, 27, 241–263.
- Poirel, L.; Héritier, C.; Tolün, V.; Nordmann, P. Emergence of Oxacillinase-Mediated Resistance to Imipenem in Klebsiella pneumoniae. Antimicrob. Agents Chemother. 2004, 48, 15–22. Mairi, A.; Pantel, A.; Sotto, A.; Lavigne, J.-P.; Touati, A. OXA-48-like carbapenemases producing Enterobacteriaceae in different niches. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 587–604.
- Poirel, L.; Héritier, C.; Tolün, V.; Nordmann, P. Emergence of Oxacillinase-Mediated Resistance to Imipenem in Klebsiella pneumoniae. Antimicrob. Agents Chemother. 2004, 48, 15–22.