Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Faisal Eudes Sam and Version 2 by Catherine Yang.
Benzothiadiazole (BTH) is a commercial chemical elicitor that can induce an innate immune response in grapevines and improve the phenolic components and color quality of grapes and corresponding products.
- benzothiadiazole
- phenolic compounds
- elicitor
- grape
- quality
1. Synthesis of Plant Phenols
The soluble form of phenolics is mainly located in the vacuoles of plant cells, which may be in free or conjugated form, while the insoluble phenolics are mainly found in the cell wall matrix [1][2][27,93]. The veraison is the critical period in grapes during which phenols develop [3][52]. Phenolic compounds are usually synthesized from phenylalanine or tyrosine through the shikimic acid pathway in plant intracellular organs during the growth of plants [1][4][5][27,64,66]. They consist of an aromatic ring with one or more hydroxyl substituents and range from monomeric molecules to highly polymerized compounds [5][6][66,94]. The hydroxyl substituents on the aromatic ring are responsible for the antioxidant properties of the phenolic compound [5][66].
The synthesis of phenolic compounds originates from the branching of phenylpropanoids [7][8][9](Figure 4) [44,62,95]. The first step of synthesis begins with the deamination of phenylalanine catalyzed by phenylalanine ammonia-lyase (PAL) [10][61]. Phenylalanine is a product of the shikimate pathway, which links carbohydrate metabolism with the production of aromatic amino acids and secondary metabolites [8][62]. In the second step, phenol skeletons are derived from malonyl-CoA and p-coumaroyl-CoA, which are biogenetically derived from phenylpropanoids and acetate pathways [8][62]. Subsequently, malonyl-CoA and p-coumaryl-CoA are transformed into phenols by stilbene synthase (STS) and chalcone synthase (CHS) through the formation of an aromatic ring (by adding three more carbon groups consisting of two C atoms) [3][7][44,52]. Through a bifurcation of this pathway, two major classes of phenolic compounds, flavonoids (by CHS) and stilbenes (by STS), can be synthesized. In addition, the flavonoid pathway leads to the synthesis of flavan-3-ols, flavonols, anthocyanins, and proanthocyanidins [8][11][12][62,96,97].
2. Influence of BTH
Presently, there are more studies on pre-harvest BTH treatment to enhance phenolic accumulation in grapes but fewer studies on the effects of BTH treatment on the phenolic metabolism of grapes. Based on the mechanism of SAR pathway, BTH (functional analog of SA) can stimulate the expression of protein defense genes (PR1, PR2, and PR5) associated with pathogenesis via the protein non-expressor gene 1 (NPR1), which is also associated with pathogenesis [13][14][15][48,98,99]. Expression of the protein defense genes then activates the SA signaling pathway leading to SAR establishment [10][14][61,98] and encoding protein defense and key enzymes of secondary metabolism (Figure 15) [3][16][17][52,59,100]. This is also accountable for the increase of phytoalexins, synthesis of protein defense genes, and reinforcement of cell walls, among others [18]. It has been reported that BTH inhibits ethylene and malondialdehyde (MDA) [19][101] but enhances the catalytic activity of various enzymes, particularly PAL, thereby influencing the synthesis of secondary metabolites in plants [20][21][22][23][24][25][10,22,50,102,103,104]. Paladines-Quezada et al. [26][30] suggested that grapevine cells treated with elicitors such as BTH activate defense responses that produce high amounts of superoxide radicals, H2O2, and other reactive oxygen species (ROS) in the cell wall. ROS can cause rapid cross-linking or interlacing of cell wall phenolic compounds. Wang et al. [21][22] also found that BTH can activate a SAR defense response in grape suspension cells and enhance the accumulation of stilbene phytoalexins, VvNPR1.1 and PR1 genes expression, and the cellular burst of H2O2. Cellular hypersensitive defense responses, including activation of defense genes and induction of defense compounds (e.g., phenols) in plants during oxidative burst could be attributed to the accumulation of H2O2 in plant cells [27][105]. Using elicitors such as BTH can induce early and rapid H2O2 production in cultured V. vinifera cells, triggering the expression of defense-related genes or phytoalexins (e.g., phenols) synthesis [21][28][29][22,106,107].
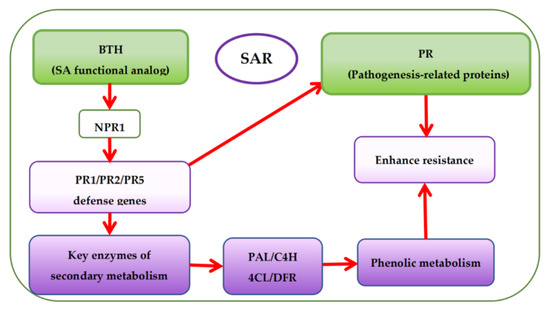
Figure 15.
Overview of systemic acquired resistance (SAR) and phenolic accumulation mechanism with BTH treatment.
However, BTH can reduce the levels of primary metabolites such as amino acids and soluble sugars in treated grapes [21][30][16,22]. In particular, total soluble sugar and soluble sugar composition may vary significantly in BTH-treated berries [21][22]. Soluble sugars are not only responsible for fruit organoleptic quality but also act as key signaling molecules that can modulate the transcription of genes involved in defense responses and metabolic processes, thus affecting the biosynthesis of secondary metabolites [21][31][22,108]. Hence, there are soluble sugars associated with BTH effect on the phenolic compounds in grapes and the defense mechanisms induced by BTH. BTH treatment can stimulate lower sucrose-synthesizing enzymes (SS-synthesis, SPS, and SPP and higher sucrose-hydrolyzing enzyme (SS-cleavage), triggering a slow increase in sucrose breakdown, a decrease in glucose level and the buildup of fructose in grapes [21][22]. In addition, sugar accumulation in grape pulp and skin can also enhance the synthesis of anthocyanins [3][32][33][52,109,110]. Therefore, soluble sugars are associated with the impact of BTH on the phenolic contents of grapes and the defense mechanisms induced by BTH. Some authors [21][22] suggest that the phenylpropanoid pathway and the common precursor of sucrose metabolism (UDP-glucose) may be directed toward the biosynthesis of phenolic compounds by BTH treatment, whereas soluble sugar accumulation might reduce.
The increase of anthocyanins and stilbenes content in BTH-treated grapes could also be due to BTH induction of PAL [34][58]. Iriti et al. [3][52] found that the buildup of stilbenes in the berry skin decreased during the ripening period of untreated berries while the production of anthocyanins increased, possibly due to competition between the two branches of the phenylpropanoid pathway and the different regulation of the key enzymes STS and CHS. Meanwhile, it appears that BTH treatment reverses the opposite association between anthocyanin and resveratrol pathways to some extent. Hence, BTH may reduce competition between STS and CHS, enabling substrate binding and increasing anthocyanin and resveratrol synthesis. In BTH-treated berries, anthocyanin accumulation does not necessarily affect resveratrol synthesis during ripening. In other words, the usual metabolic switch between the two branches of the same metabolic pathway seems to be avoided by BTH treatment [3][52].
Anthocyanins can be affected by BTH treatments [35][36][23,111]. Gómez-Plaza et al. [35][23] demonstrated that BTH could activate enzymes related to anthocyanin metabolism. Repka et al. [37][112] and Gozzo [38][113] also reported that BTH treatment promoted the activities of enzymes (i.e., chalcone isomerase and PAL) in the phenylpropanoid pathway. Furthermore, BTH treatment can contribute to stilbenes and flavan-3-ols synthesis due to SAR and BTH induction of the expression of phenylpropanoid genes in grapevine [8][62]. In addition, in light of the effect of BTH treatment on grape polyphenols and the findings of earlier studies, Fumagalli et al. [16][59] suggested that BTH could enhance the activity of CHS during polyphenol or anthocyanin biosynthesis.
Studies on pre- or post-harvest BTH treatment have been conducted not so much in terms of secondary metabolic mechanisms in grapes as in various plants. In one study, post-harvest BTH treatment resulted in a higher anthocyanin content in strawberries and increased the activities of metabolic enzymes such as PAL, 4-coumarate/coenzyme A ligase (4-CL), cinnamate-4-hydroxylase (C4H), dihydroflavonol 4-reductase (DFR), glucose-6-phosphate dehydrogenase (G6PDH), tyrosine ammonia-lyase (TAL), and shikimate dehydrogenase (SKDH) [36][111]. This suggests that the increase in anthocyanin content by BTH may be due to the activation of associated metabolic enzymes. In addition, post-harvest BTH treatment resulted in the activation of PAL in peaches and mangoes [39][40][114,115], while the activity of peroxidases (POD) and polyphenol oxidase (PPO) increased in mangoes [40][115]. Similarly, post-harvest BTH treatments also induced PPO and POD activity in bananas [41][116] and loquats [42][117]. Interestingly, this approach has been reported to increase the total phenolic compounds in mangoes and bananas [40][41][115,116]. Furthermore, the exogenous application of BTH increases the expression of proanthocyanidin-related MBW complex (MYB-bHLH-WD40) and the content of flavan-3-ol in general [43][118], which promotes the accumulation of proanthocyanidins. Similar results were also reported by Felicijan et al. [44][119]. However, the activity and gene expression of phospholipase (phospholipase A2PLA2, phospholipase CPLC, and phospholipase DPLD) were inhibited by post-harvest BTH treatment in melons [45][54].
Overall, the influence of BTH on grapes has been reported in the literature, particularly on grape color, phenolics, variety, and metabolism (Table 1). Based on the existing literature, many studies have been conducted on the effects of BTH on grape phenolics in relation to variety, but less on the effects on grape metabolism and sensory characteristic such as color and mouthfeel (which are associated with phenolic compounds). Hence, future studies can be conducted in these areas.
Table 1.
Summary of literature on BTH impact on grape variety, phenols, color, and metabolism.
| Research Area | Subclasses | Reference |
|---|---|---|
| Variety | Monastrell | [7][10][26][35][46][47][48][49][50][51][23,30,33,44,46,47,61,82,87,88] |
| Merlot | [3][10][20][35][48][51][10,23,47,52,61,88] | |
| Syrah | [10][35][51][52][53][23,49,53,61,88] | |
| Cabernet Sauvignon | [48][49][47,82] | |
| Groppello | [54][2] | |
| Beauty Seedless | [55][83] | |
| Cabernet Gernischt | [30][16] | |
| Phenols | Anthocyanin | [3][7][10][20][26][35][46][47][10,23,30,33,44,46,52,61] |
| Flavonols | [7][35][46][47][50][51][23,33,44,46,87,88] | |
| Proanthocyanidins (Tannins) | [10][22][35][51][1020],23[,50,61,88] | |
| Stilbenes | [3][21][50][53][22,52,53,87] | |
| Metabolism | Enzymes | [3][20][21][22][35][10,22,23,50,52] |
| Hydrogen dioxide and ROS | [26][35][23,30] | |
| Color | Color intensity | [7]16[10][30][35][48][51][,23,44,47,61,88] |