2. Transcription of Drought-Responsive Genes
2.1. Drought-Responsive Genes
More than half of the genes up- or downregulated under drought or high-salinity conditions are also regulated by ABA application, suggesting that the expression of osmotic stress-responsive genes is driven mainly by ABA
[6][7][31][6,7,34]. ABA induces the expression of numerous genes encoding enzymes that catalyze the biosynthesis of osmoprotectants such as trehalose and late embryogenesis-abundant proteins. In addition, ABA-induced genes include those encoding proteins exerting either positive or negative effects on its accumulation (biosynthesis, catabolism, and glucose-conjugation), transportation, and signaling network
[32][35]. By analyzing the promoters of ABA-responsive genes, a conserved
cis-acting ABA-responsive element (ABRE; PyACGTGG/TC) was identified
[33][36]. Subsequently, several ABRE-binding (AREB) proteins and ABRE-binding factors (ABFs) were identified by yeast one-hybrid screening
[34][35][37,38].
However, a few drought-inducible genes do not respond to ABA, implying that ABA-independent pathways regulate the drought response
[36][39]. The promoters of these genes contain the
cis-acting element DRE (dehydration-responsive element)/CRT (C-repeat), which functions in ABA-independent gene expression
[33][36]. The ERF/AP2 family transcription factors CBF/DREB1 (C-repeat-binding factor/DRE binding 1) and DREB2, which bind to DRE/CRT elements, were identified in plants
[33][36]. Most CBF/DREB1 target genes in Arabidopsis (
Arabidopsis thaliana) contain the DRE motif with a conserved (A/G)CCGACNT sequence in their promoter regions. Rice (
Oryza sativa) genome sequence analyses identified 10
OsDREB1s and 4
OsDREB2s, indicating that similar transcription factors function in drought stress tolerance in dicotyledonous and monocotyledonous plants. The Arabidopsis
RD29A (
responsive to desiccation 29A) contains both
cis-acting elements, ABRE and DRE/CRT, in the promoter region.
2.2. Regulation of ABA Signaling
The ABA de novo synthesis pathway has been unraveled
[4][37][4,40]. ABA is biosynthesized in vascular tissues and transported to targets including guard cells via the export and import of transporters. In Arabidopsis, three transporter families have been identified: ATP binding cassette (ABC), detoxification efflux carriers (DTX)/multidrug and toxic compound extrusion (MATE) proteins, and nitrate transporter 1/peptide transporter family (NPF)
[32][38][39][40][35,41,42,43]. For instance, ABCG25 exports ABA to the apoplastic space in vascular tissues, while ABCG40 and NPF4.6 import ABA into guard cells in response to osmotic stress.
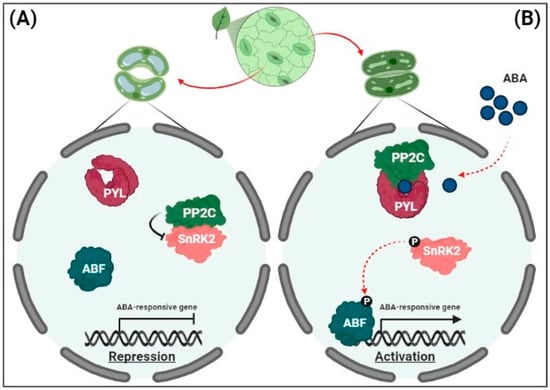
In target cells, ABA is perceived by soluble receptors in the nucleus and cytosol. Several synonymous ABA receptors, i.e., PYR (pyrabactin resistance), PYL (PYR-related), and RCAR (regulatory component of the ABA receptor), have been identified in Arabidopsis
[41][42][44,45]. In the absence of ABA under non-stressful conditions, clade A type 2C protein phosphatases (PP2Cs) in guard cells counteract a family of protein kinases, known as sucrose non-fermenting 1-related protein kinase 2s (SnRK2s) via physical interactions, thus providing negative feedback regulation of ABA signaling
[43][44][46,47] (
Figure 1A). Under osmotic stress, following ABA perception, PP2Cs bind to ABA receptors to capture ABA and form the PYL-ABA-PP2C complex; SnRK2s subsequently dissociate from inactivated PP2Cs to restore kinase activity
[45][48] (
Figure 1B).
Figure 1. Signaling pathway for the expression of abscisic acid (ABA)-responsive genes. (A) Repression of ABA-responsive genes. In the absence of ABA, clade A protein phosphatases (PP2Cs) physically interact with sucrose non-fermenting 1-related protein kinase 2s (SnRK2s) to reduce kinase activity via dephosphorylation. This results in the inhibition of ABRE-binding (AREB)/ABRE-binding factors (ABFs) and the suppression of ABA-responsive gene transcription. (B) The activation of ABA-responsive gene expression. Under drought stress, soluble ABA receptors (Pyrabactin resistance/PYR-related/regulatory component of the ABA receptors [PYR/PYL/RCARs]) and PP2Cs act as co-receptors to capture ABA, thereby blocking the phosphatase activity of PP2Cs. PP2Cs are released from PP2C-SnRK2 complexes, and free SnRK2s phosphorylate downstream transcription factors (AREBs/ABFs). The phosphorylated AREBs/ABFs trigger the transcription of numerous ABA-responsive genes, leading to ABA responses including stomatal closure.
Activated SnRK2s in the cytosol facilitate the functioning of slow anion channel 1 (SLAC1), K
+ channel 1 (KAT1), and NADPH oxidases in the guard cell membrane to induce stomatal closure
[32][46][35,49]. In the nucleus, under osmotic stress, activated SnRK2s phosphorylate and activate a family of basic-domain leucine zipper (bZIP) transcription factors, the AREB/ABFs, thereby inducing the expression of numerous ABA-responsive genes
[47][50]. Among the nine AREB/ABFs in Arabidopsis, ABF1, AREB1/ABF2, AREB2/ABF4, and ABF3 act as master transcription factors in ABA signaling to promote osmotic stress tolerance
[48][51].
3. Transcriptional Memory of Drought Tolerance
3.1. Drought Stress Memory
Plants memorize the tolerance induced by drought stress to respond more effectively to subsequent stresses
[13][49][50][13,149,150]. For instance, Walter et al.
[51][151] observed that after a late drought during the growth of perennial grass (
Arrhenatherum elatius), the percentage of living biomass was increased in plants exposed to earlier drought compared to those without such exposure, even after harvest and resprouting after the first drought. Ding et al.
[52][152] reported that Arabidopsis with experience of dehydration stress wilted more slowly than plants without such experience in response to subsequent dehydration events. Wang et al.
[53][153] showed that wheat plants subjected to one or two drought episodes before anthesis had higher grain yields under drought conditions. Ramírez et al.
[54][154] reported that long-term stress improved drought tolerance-related traits and tuber yield in later growth stages in potato plants. Abdallah et al.
[55][155] showed that the pre-exposure to a drought-sensitive variety of olive plants to drought enhanced their tolerance to subsequent drought conditions, resulting in improvements in biomass production, photosynthesis, and the maintenance of water status. Tabassum et al.
[56][156] also reported that terminal drought and seed priming improved the drought tolerance of wheat plants.
Goh et al.
[57][157] observed that Arabidopsis exhibited memory functions related to repeated ABA stresses, i.e., the impairment of light-induced stomatal opening and the induction of the expression of drought-responsive genes. Virlouvet and Fromm
[58][158] reported that Arabidopsis stomatal apertures closed following exposure to dehydration remained partially closed during a recovery period with access to water, thereby facilitating reduced transpiration during subsequent dehydration stress. In mutant plants defective in the ABA signaling pathway, the guard cell stomatal memory was ABA-dependent, and
SnRK2s were essential for implementing stress memory during the subsequent dehydration response. Li et al.
[59][159] performed whole-transcriptome, strand-specific RNA sequencing (ssRNA-seq) of the rice genome, and the results suggested that lncRNAs, DNA methylation, and endogenous ABA mediate drought memory by activating the drought-responsive transcription of genes in pathways such as photosynthesis and proline biosynthesis in response to subsequent drought conditions.
3.2. Drought Stress Memory Genes
In response to drought stress, plants induce or suppress the expression of many drought-responsive genes
[1][2][1,2]. In most cases, up- or downregulated gene transcripts return to basal levels during recovery (watered) states. However, a subset of genes is expressed at highly elevated or reduced levels in response to repeated drought stresses, which enables the plant to respond more promptly and strongly
[13][16][52][13,16,152]. Ding et al.
[60][160] defined stress memory genes as those that enable responses during subsequent stress conditions that differ from the responses during the initial stress encounter, whereas genes that respond similarly to each stress are categorized as non-memory genes. However, the expression threshold for classification into one or the other category is unclear.
Numerous drought stress memory genes have been identified in plants. Ding et al.
[60][160] used a genome-wide RNA sequencing (RNA-seq) approach to evaluate the transcriptional responses of Arabidopsis leaves detached from plants repeatedly exposed to air-drying. Genes implicated in the responses to ABA, drought, salinity, and cold/heat acclimation constituted the drought-induced memory genes, and those responsible for chloroplast and thylakoid membrane-associated functions comprised the dehydration-repressed memory genes. Kim et al.
[61][161] performed a microarray analysis to screen drought stress memory genes in soybean. The soybean memory genes exhibiting significantly elevated transcript levels upon the second exposure to drought stress conditions include those involved in ABA-mediated tolerance responses to abiotic stresses, such as genes encoding transcription factors, trehalose biosynthesis enzymes, late embryogenesis abundant proteins, and PP2C family proteins. By contrast, memory genes with highly reduced transcript levels during the second drought included genes involved in photosynthesis and primary metabolism. Soybean drought stress memory genes included genes involved in the dehydration memory responses of Arabidopsis. However, studies of other crop plants identified species-specific drought memory genes. A genome-wide RNA-seq analysis of maize identified only 4 chloroplast- and 2 thylakoid membrane-localized genes acting as drought-repressed memory genes
[62][162], compared to 128 Arabidopsis drought-repressed memory genes
[60][160]. In potato, the expression levels of most photosynthesis-related genes during a second drought were higher than during the first drought
[63][163]. In addition, most rice memory transcripts associated with photosynthesis were markedly reduced by a first drought but then recovered, remaining at a stable level during subsequent drought treatments
[59][159].
Several memory genes encode various transcription factors in all of the plant species described above. In Arabidopsis, dehydration-induced transcriptional memory behavior was seen in members of the AP2/ERF, bHLH, homeo_ZIP, MYB, ZF, b_ZIP, CCAAT, and WRKY transcription factor families
[60][160]. Similarly, in soybean, various transcription factor genes belonging to the AP2, NAM, MYB, bZIP_1, and WRKY families were identified as drought-induced memory genes
[61][161]. Therefore, transcription factors with memory function may contribute to plant memory and regulate the expression of their targets upon repeated stress. This possibility should be addressed in further studies of the mechanisms of drought stress memory.
3.3. Mechanism of Transcriptional Stress Memory
3.3.1. Epigenetic Marks for Stress Memory
Epigenetic chromatin remodeling is a plausible molecular mechanism of transcriptional stress memory
[12][22][23][50][12,22,23,150]. It has been proposed that chromatin architecture (DNA methylation, histone modification, and chromatin loops) maintains the altered gene expression patterns caused by an initial stressor. Transcription factors and chromatin loops may also be associated with a subset of their targets during mitosis. The role of the chromatin remodeler BRM in stress memory under heat-shock stress has been investigated. Brzezinka et al.
[64][164] reported that Arabidopsis
brm mutants were deficient in heat-shock memory and showed the reduced induction of heat-shock memory genes. BRM and the FORGETTER1 (FGT1) factor physically interact and are pre-associated with memory genes under non-stress conditions.
fgt1 mutants displayed more rapid recovery of nucleosome occupancy at heat-shock memory gene loci, suggesting that the BRM-FGT1 interaction prevents nucleosome recovery at these loci and mediates heat stress-induced memory.
Changes in DNA methylation may be involved in the transcriptional memory of plant responses to abiotic stresses
[65][72]. Wang et al.
[66][63] observed that around 29% of drought-induced DNA (de)methylation sites remained after recovery from drought or salt stress, implying that the DNA methylation changes were recorded. Kou et al.
[67][165] performed a genome-wide rice methylome profiling analysis under recurrent drought stresses and recovery treatments. Most drought-stress memory-related DMRs were targeted TEs and few were targeted gene bodies, which suggests that they regulate TE expression to cope with recurrent drought stress. The distances from memory DMRs to TEs were significantly shorter than those from non-memory DMRs, implicating DNA methylation in drought memory formation.
Histone modification may provide a persistent epigenetic transmission mechanism associated with transcriptional memory in response to osmotic stress
[18][20][21][18,20,21]. For instance, H3K27me3 is a gene silencing mark related to the chromatin-induced repression of gene expression and the formation of an epigenetic memory system during development
[68][166]. In eukaryotes, the H3K27 methylation level is regulated by the action of polycomb group (PcG) protein complexes. Polycomb-repressive complex 2 (PRC2) mediates the deposition of H3K27me2/3 by the enzymatic subunit PRC2-Ezh2 (enhancer of ZESTE 2), whereas PRC2-Ezh1 restores H3K27me2/3 via its demethylase activity or histone exchange
[69][167]. Ezh1 and Ezh2 exhibit different expression patterns and distinct chromatin-binding properties
[70][168]. H3K27me3 in target genes recruits an additional PcG protein complex, PRC1. PRC1 complexes are subdivided into canonical PRC1 and noncanonical (or variant) PRC1. Canonical PRC1 is recruited by H3K27me3 readers and compacts nucleosomes to repress gene expression, while noncanonical PRC1 is recruited to chromatin independently of PRC2 and H3K27me3, and ubiquitylates histone H2A (to form H2AK119ub) via its H2A E3 ubiquitin ligase activity
[71][72][169,170]. Thus, PcG complexes provide the major chromatin regulatory mechanism for silencing unnecessary or unwanted gene expression in mammals and plants
[73][74][171,172].
PcG genes were discovered in
Drosophila (
Drosophila melanogaster), and homologs of PcG components and their target genes have been identified in other eukaryotes including plants
[75][76][77][78][173,174,175,176]. The role of the PRC2-mediated deposition of H3K27me3 has been studied in the context of developmental processes and environmental stress responses in plant model species including Arabidopsis
[74][79][80][81][172,177,178,179]. The PRC1-like protein LHP1 (like heterochromatin protein-1) was also identified in Arabidopsis
[68][82][83][166,180,181]. Ramirez-Prado et al.
[84][182] showed that the loss of
LHP1 induces ABA sensitivity and drought tolerance, indicating that LHP1 regulates the expression of stress-responsive genes. The H3K27me3 level was not related to H3K4me3 accumulation, suggesting that these histone modification marks function independently and do not have mutual effects on the expression of dehydration stress memory genes.
H3K4me3 deposition may play a role in the epigenetic transmission of active transcriptional states
[19]. H3K4me3 deposition and the amount of Pol II stalled in memory genes were higher than in non-memory genes in Arabidopsis after multiple dehydration stresses
[52][85][86][152,183,184]. Kim et al.
[87][185] observed H3K4me3 and H3K9ac deposition in drought-inducible genes (
RD20,
RD29A, and
AtGOLS2) in response to drought. During recovery by rehydration, H3K9ac was rapidly removed, whereas H3K4me3 was maintained at a low level, implying that H3K4me3 functions as an epigenetic mark of stress. Ding et al.
[52][152] observed that H3K4me3 deposition in trainable genes (
RD29B and
RAB18) was maintained during recovery from stress-induced transcription but decreased to a basal level in non-trainable genes (
RD29A and
COR15A). Thus, H3K4me3 and Pol II were induced in several dehydration stress memory genes in response to the first dehydration stress event, persisted during the recovery period, and increased greatly due to a second stress event.
As identified in
Drosophila, the epigenetic transmission of active transcriptional states is supposed to be mediated by TrxG (trithorax group) complexes: the SWI/SNF complex and the COMPASS (complex of proteins associated with Set1) family
[88][89][186,187]. Antagonistic links were identified between PcG genes and SWI/SNF, and COMPASS was associated with histone methyltransferase activity leading to the H3K4me3 deposition. In Arabidopsis, multiple TrxG factors have been identified, based on their ability to suppress PcG mutant phenotypes
[90][188]. Plant TrxG factors regulate gene transcription in seedling growth, anther and ovule formation, gametophyte development, and reprogramming during developmental transitions
[91][92][93][94][189,190,191,192].
However, it is not clear whether H3K4me3 in chromatin contributes to transcriptional activation under subsequent stress conditions. Genome-wide transcript profiling revealed that the transcription of most genes is unaffected by the loss of the histone methyltransferase activity of ATX1 (Arabidopsis homolog of TRITHORAX 1) and that H3K4me3 is required for efficient elongation of the transcription, but not the initiation, of ATX1-regulated genes
[95][96][193,194]. Howe et al.
[97][195] proposed that H3K4me3 deposition in chromatin is a consequence of transcription, influencing splicing, transcription termination, the memory of previous states, and transcriptional consistency, rather than inducing gene transcription in response to repeated stresses. Moreover, the mechanism of TrxG recruitment to the chromatin during mitosis is not clearly elucidated.
4. Transgenerational Inheritance of Memory
4.1. Transgenerational Transmission of Drought Tolerance
Traits acquired under stressful conditions can be transmitted to plant progeny
[29][30][98][29,30,202]. The progeny of parents exposed to stress exhibits a higher yield than the progeny of non-stressed parents. Lämke and Bäurle
[21] use the term ‘intergenerational memory’ when only the first stress-free generation has a detectable memory effect. As the progeny develops on the mother plant, intergenerational memory may be mediated by the conditions in which the seed grows and by cues introduced into the seed or embryo by the mother plant. In ‘transgenerational memory’, by contrast, memory is detectable after at least two stress-free generations. Verkest et al.
[99][203] improved drought tolerance in canola (
Brassica napus) by repeatedly selecting lines exhibiting increased drought tolerance for three generations. Tabassum et al.
[100][204] reported that the hydro- and osmo-priming of bread wheat seeds caused the transgenerational transmission of improved tolerance to drought and salt stresses. Raju et al.
[101][205] applied RNAi suppression to modulate abiotic stress-response pathways in soybean and developed an epigenetic breeding system for increased yield and stability.
Transgenerational memory may have an epigenetic basis
[21]. Based on invertebrate developmental processes, it has been suggested that histones and other core chromatin components survive the passage of replication forks during meiosis
[102][103][104][206,207,208]. Zenk et al.
[105][209] showed that H3K27me3 was transgenerationally inherited from the maternal germline and resisted reprogramming events, thereby regulating the activation of enhancers and lineage-specific genes during early embryogenesis in
Drosophila. Molla-Herman et al.
[106][210] proposed that chromatin modifiers and Piwi-interacting small RNAs (piRNAs) function in adaptive and inheritable epigenetic memory events that occur in
Drosophila during embryogenesis. Weiser and Kim
[107][211] revealed an important role of endogenous siRNAs and piRNAs in transgenerational epigenetic inheritance in
Caenorhabditis elegans. Those sRNAs may regulate heritable chromatin marks conveying epigenetic memory and thereby repress deleterious transcripts, such as TEs and repetitive elements. However, the relevance of these mechanisms to plant transgenerational inheritance of drought stress tolerance is unclear.
4.2. DNA Methylation for Transgenerational Inheritance
DNA methylation may contribute to transgenerational memory in plants
[65][72]. Zheng et al.
[108][212] reported that a high proportion of multigenerational drought-induced alterations in DNA methylation status are maintained in subsequent generations, possibly improving drought adaptability in rice. Drought stress-induced non-random epimutations over 11 successive generations improved the drought adaptability of rice epimutation lines. A large proportion (~45%) of the altered DNA methylation states were transmitted to unstressed progeny in subsequent generations. The epimutated genes participated in stress-responsive pathways, suggesting that they promote progeny adaptation to drought stress. Mathieu et al.
[109][213] observed that the Arabidopsis mutant
methyltransferase 1-3 (
met 1-3), which is deficient in terms of maintaining CG methylation, formed progressively more aberrant epigenetic patterns over several generations, suggesting that CG methylation is a central coordinator of epigenetic memory that secures stable transgenerational inheritance. Zhang et al.
[110][214] reported that many epigenetic recombinant inbred lines of Arabidopsis were nearly isogenic as a result of drought but were highly variable at the level of DNA methylation. Cortijo et al.
[111][215] identified several DMRs that act as epigenetic quantitative trait loci and account for 60–90% of the heritability related to flowering time and primary root length in Arabidopsis. However, the contributions of locus-specific methylation changes to the maintenance of stress memory and whether the inheritance of drought stress tolerance is mediated only by DMRs require further investigation.
Matzke and Mosher
[112][216] proposed that RdDM contributes to the transmission of DNA methylation patterns in parental cells to their offspring by affecting germ-cell specification and parent-specific gene expression. Morgado et al.
[113][217] observed that the composition of sRNAs in apomictic dandelion (
Taraxacum officinale) lineages indicated a footprint of drought stress experienced two generations prior. Kuhlmann et al.
[114][218] reported that the methylation of the reporter gene ProNOS was not completely erased in
DRM-2 (
domains rearranged methyltransferase 2) mutants but persisted in the context of symmetric CG. ProNOS DNA methylation maintenance was evident after two generations of ongoing RdDM and increased in subsequent generations. They suggested that the methylation of a particular genomic region can be consolidated by RdDM and maintained over generations in Arabidopsis, thereby establishing epigenetic transgenerational memory. Wibowo et al.
[115][67] suggested that epigenetic inheritance relies on DNA methylation changes at sequences that function as distantly acting control elements of key stress-response regulators, including antisense lncRNAs. Some of these changes are associated with conditionally heritable adaptive phenotypic stress responses and transmitted to the offspring, where they affect the transcriptional regulation of a small group of genes associated with enhanced tolerance to environmental stresses.
By contrast, two studies of Arabidopsis yielded conflicting results. In Arabidopsis subjected to slow-onset water deprivation treatment, Ganguly et al.
[116][219] observed far fewer conserved DMRs in drought-exposed lineages compared to non-exposed lineages. Most of the variation was attributed to preexisting differences in the epigenome at repetitive regions of the genome. Thus, transgenerational memory may not be associated with changes in the DNA methylome. Van Dooren et al.
[117][61] found that descendants of stressed and non-stressed Arabidopsis plants were phenotypically indistinguishable after an intervening generation without stress, irrespective of whether they were grown under normal or water-deficit conditions. In addition, although mild drought induced changes in the DNA methylome of exposed plants, these were not inherited by the next generation. Therefore, whether stress-induced DNA methylation variation transmits drought stress memory to the next generation is unclear.
4.3. Overcoming Meiosis
In mammals, paternal chromatin is extensively reprogrammed via the global erasure of DNA methylation achieved through extensive DNA demethylation and the packaging by exchange of histones with protamines
[118][119][220,221]; this hampers the inheritance of stress-induced changes in chromatin architecture. Thus, epigenetic marks are reprogrammed in the gametes and the genomic potential is thus reset in the next generation. In contrast to mammals, DNA methylation in flowering plants is not completely erased from the germlines and is thus maintained during reproduction
[120][121][122][222,223,224]. Wibowo et al.
[115][67] reported that hyperosmotic stress memory in Arabidopsis restricts DME activity in the male germline. Moreover, protamine exchange does not occur in Arabidopsis, enabling the retention of histone-based chromatin in sperm
[123][225]. However, Borg et al.
[124][226] found that H3K27me3 is completely lost from histone-based sperm chromatin in Arabidopsis by the concerted action of three mechanisms; (1) the loss of histone methyltransferase activity in PRC2 to write H3K27me3, (2) the erasing activity of H3K27 demethylases, and (3) the deposition of the sperm-specific histone variant H3.10 which may be resistant to H3K27 methylation. The loss of H3K27me3 facilitates the transcription of genes essential for spermatogenesis but resets epigenetic memory in plant paternal chromatin.
Based on the above observations, newly acquired stress tolerance and associated epigenetic marks might be preferentially transmitted through the female germline. In Arabidopsis, Borg et al.
[124][226] detected H3K27me3 in the microspore and in the daughter nuclei following microspore division, suggesting inheritance via meiosis. Inoue et al.
[125][227] identified maternal H3K27me3 as a DNA methylation-independent imprinting mechanism in mouse (
Mus musculus) embryonic cell lineage. Grossniklaus and Paro
[126][228] reported that no major loss of H3K27me3 is expected on maternal alleles because PRC2 is active in the central cell. PcG complexes deposit or bind to certain histone modifications (e.g., H3K27me3 and H2AK119ub1) to prevent gene activation and maintain the repression of chromatin domains, which are implicated in plant vernalization and seed development. The relevance to plant drought stress tolerance mechanisms identified in flowering warrants further investigation.