Xmrk is a gene product closely related to the human epidermal growth factor receptor (EGFR), which is associated with a wide variety of pathological conditions, including cancer. Comparative analyses of Xmrk and EGFR signal transduction in melanoma have shown that both utilize signal transducer and activator of transcription 5 (STAT5) signaling to regulate apoptosis and cell proliferation, phosphoinositide 3-kinase (PI3K) PI3K to modulate apoptosis, focal adhesion kinase (FAK)FAK to control migration, and the Ras/Raf/MEK/mitogen-activated protein kinase (MAPK)MAPK pathway to regulate cell survival, proliferation, and differentiation. Further, Xmrk and EGFR may also modulate similar chemokine, extracellular matrix, oxidative stress, and microRNA signaling pathways in melanoma. In hepatocellular carcinoma (HCC), Xmrk and EGFR signaling utilize STAT5 to regulate cell proliferation, and Xmrk may signal through PI3K and FasR to modulate apoptosis. At the same time, both activate the Ras/Raf/MEK/MAPK pathway to regulate cell proliferation and E-cadherin signaling.
1. Introduction
The epidermal growth factor receptor (EGFR) is a member of the erythroblastic leukemia viral oncogene (ErbB) family of receptor tyrosine kinases (RTKs). Ligand-dependent EGFR activation triggers receptor autophosphorylation creating phosphotyrosine-binding sites for other proteins that then signal along various signal transduction pathways, e.g., mitogen-activated protein kinase (MAPK), phosphoinositide 3-kinase/protein kinase B (PI3K/Akt) and c-Jun N-terminal kinase (JNK), which control cellular behaviors, including adhesion, migration and proliferation
[1][2][1,2] ( and ). Mutations in EGFR have been linked to developing various diseases, including diverse cancers, including melanoma and hepatocarcinoma (HCC)
[3][4][3,4]. Comparative genomic studies have shown that species of many other organisms, including fish, have genetic sequences homologous to EGFR
[5].
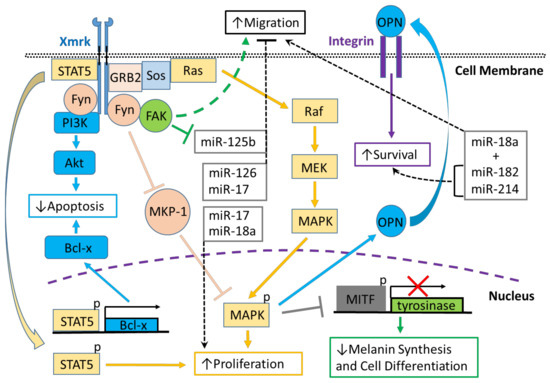
Figure 1. Xmrk signaling in the Xiphophorus melanoma model. Xiphophorus melanoma receptor kinase (Xmrk) is a receptor tyrosine kinase that recruits and activates several kinases, transcription factors and adaptor proteins integrated into signaling networks to regulate pigment cell cancer transformation. Activation of the kinase, phosphoinositide 3-kinase (PI3K), prevents apoptosis in malignant cells, as does activation of the transcription factor, signal transducer and activator of transcription 5 (STAT5), via B-cell lymphoma (Bcl-x) signaling, and STAT5 also promotes proliferation. The Src kinase protein, Fyn, acts as a docking protein and, through focal adhesion kinase (FAK), can induce pigment cell migration. Further, Fyn inhibits MAPK phosphatase 1 (MKP-1), a repressor of MAPK, causing the promotion of MAPK’s function to increase melanoma cell proliferation. Xmrk can also signal via the adaptor proteins, GRB2 and Sos, to activate the Ras/Raf/MEK/MAPK pathway, thereby enhancing proliferation. Additionally, MAPK signaling causes degradation of the transcription factor, microphthalmia transcription factor (MITF), leading to reduced transcription of tyrosinase, which decreases melanin synthesis and inhibits pigment cell differentiation. Another function of activated MAPK is to induce the synthesis of osteopontin (OPN), which is secreted, binds to integrins on the cell membrane surface, and promotes pigment cell survival. Several miRNAs that are regulated in fish Xmrk models also have functions in human melanoma. The miRNAs miR-18a, -182, and -214, can act to promote migration in human melanoma, while miR-182 and -214 can also stimulate cell survival. MiR-17, 125b, and -126 act to inhibit melanoma cell migration, with -125b’s action inhibited by FAK. MiR-17, along with miR-18a, also function to increase melanoma proliferation.
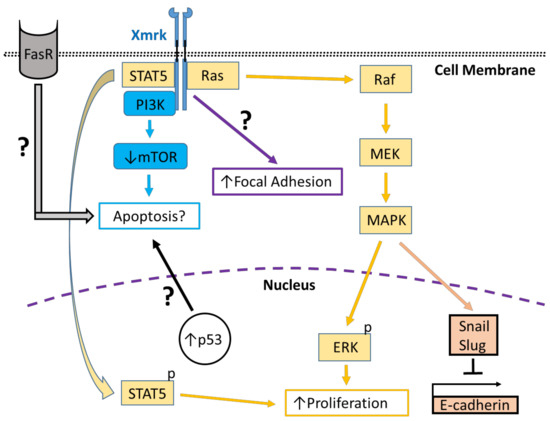
Figure 2. Xmrk signaling in the zebrafish hepatocellular carcinoma model. In transgenic zebrafish models of HCC, Xmrk signals through kinases and transcription factors that modulate liver cancer progression. PI3K activation may modulate apoptosis through the mechanistic target of rapamycin (mTOR), although mTOR expression may be decreased in HCC, making its effect on apoptosis uncertain. STAT5 signaling can promote proliferation, as does MAPK via activation of extracellular signal-regulated kinases (ERK). MAPK also has a dual role in signaling through the transcription factors, Snail and Slug, to inhibit E-cadherin activity. The formation of focal adhesions is increased through a pathway that is currently uncharacterized. Xmrk is also associated with increased expression of the tumor suppressor, p53, and the Fas receptor (FasR), which regulates apoptosis during HCC, but the details of their associated signaling mechanisms are not currently understood.
Crosses between species of the fish genus
Xiphophorus:
X. maculatus (the southern platyfish) and
X. hellerii (the green swordtail) produce fish that spontaneously develop malignant melanoma induced by a proto-oncogene encoding a receptor tyrosine kinase designated
Xiphophorus melanoma receptor kinase (Xmrk), which is located on the sex chromosome
[6]. The encoded Xmrk protein is structurally related to the human EGFR with an extracellular ligand-binding domain, a transmembrane domain and an intracellular catalytic domain
[7]. Studies in human melanoma cell lines and the
Xiphophorus cross-derived melanoma cell line (PSM-1)
[8] using chimeric receptors combining the human epidermal growth factor receptor (HER) with Xmrk (HER-mrk) have shown that Xmrk displays constitutive autophosphorylation and is a functional receptor tyrosine kinase with activity in malignant melanoma
[9]. Blockage of Xmrk’s kinase activity in
Xiphophorus derived PSM cells demonstrates that receptor autophosphorylation is sufficient to trigger mitogenic signaling in melanoma cells
[10]. Further, receptor stimulation induces autophosphorylation resulting in the production of the physiological ligand in an autocrine manner
[11].
The role of Xmrk in melanoma also has been investigated in
Oryzias latipes, the medaka fish. Ectopic expression of the Xmrk oncogene cloned from
Xiphophorus and introduced into medaka embryos induces tumors, typically classifiable as cysts or hyperplasias, primarily in the epithelium, but also in the brain, retina, integument and pigment cells
[12]. Expression of HER-mrk in medaka has demonstrated that one ligand, human transforming growth factor-alpha (hTGF alpha), can cause activation of this chimeric receptor
[12]. Mutational studies of Xmrk in medaka embryos also have shown that point mutations can lead to constitutive, ligand-independent receptor kinase activity
[13]. Medaka expressing HER-mrk, which exhibits invasive melanoma, has been used to show that activated Xmrk induces EGFR expression and ligand production leading to the formation of an autocrine loop that promotes pro-tumorigenic signaling
[14].
RNA-Seq comparative analyses of
Xiphophorus hellerii/maculatus hybrid fish and human melanoma tissues have shown shared pathways in these tissues governing: inflammation, cell migration, cell proliferation, pigmentation, cancer development, and metastasis
[15]. Structural studies using the
Xiphophorus and medaka models suggest that oncogenic signaling mechanisms and related translational studies can be studied using zebrafish transgenic Xmrk models. Recently, the tetracycline-controlled transcriptional activation Tet-on system
[16][17][16,17] has been used in zebrafish to express Xmrk under the control of the liver-specific fatty acid-binding protein 10 (fabp10) promoter (also designated as liver fatty acid-binding protein (LFABP)) to study signaling in hepatocellular carcinoma (HCC)
[18][19][20][21][18,19,20,21].
2. Genetic Characterization and Comparison of Xmrk with the Human EGFR
The Xmrk gene is hypothesized to have arisen from gene duplication and nonhomologous recombination during evolution
[5][22][5,22]. There are three
Xiphophorus orthologs of the EGFR gene: egfra, which encodes a tyrosine kinase receptor; egfrb, which is a proto-oncogene form termed INV-Xmrk; and ONC-Xmrk, a duplicated version of INV-Xmrk, which is highly expressed in melanoma where it is responsible for inducing oncogenic transformation
[23][24][25][26][27][23,24,25,26,27]. Oncogenic activation of the ONC-Xmrk receptor occurs via two mutations, i.e., G336R and C555S, both located in the extracellular domain that is not found in INV-Xmrk; these mutations result in constitutive receptor dimerization leading to constitutive kinase signaling
[27][28][27,28].
The Xmrk promoter is highly methylated in nontransformed tissues, but in the
Xiphophorus cross-derived melanoma cell line (PSM-1)
[8], it is unmethylated, suggesting that methylation status also regulates Xmrk expression
[29]. Transcription of Xmrk does not stem from the action of cis-regulatory elements present in the promoter region of pigment cells, suggesting that epigenetic mechanisms also play a role in Xmrk’s oncogenic activation
[30].
In comparison to Xmrk, the human EGFR is encoded by a single gene that has orthologs in all vertebrate species
[5][31][5,31]; two transcriptions start sites have been identified, resulting in two major transcripts (10 and 5.6 kb) encoding the full-length receptor
[31]. In addition to the two transcripts that encode EGFR Isoform A (the “full-length” or holo-receptor), three alternately processed transcripts encode alternate EGFR receptor isoforms (isoforms B, C, and D)
[31], all of which are expressed in normal human tissues. While the full-length EGFR isoform (A) is known to signal through both ligand-dependent and independent signaling pathways
[32], all three alternate isoforms lack the receptor kinase domain, rendering these isoforms unable to signal directly through protein phosphorylation. These isoforms have been hypothesized to function through signaling mechanisms similar to those used by the IGF-binding proteins, including regulating cell surface to extracellular matrix interactions, i.e., as matricellular proteins
[33].
The transmembrane glycoprotein EGFR is derived from the 1210 residue precursor, which is cleaved at the N-terminus resulting in an 1186 residue mature EGFR protein
[34]. The extracellular or ligand-binding domain (exons 1–16, amino acids 1–621) of the human EGFR receptor comprises four subdomains, two leucine-rich repeats (I and III) and two cysteine-rich repeats (II and IV)
[35]. Subdomains I and III are involved in ligand-binding, domain II facilitates the formation of homo- or heterodimers with an analogous family member domain, and domain IV forms disulfide bonds to domain II. These subdomains are followed by a single hydrophobic transmembrane domain (exon 17, amino acids 645–668) and an intracellular cytoplasmic domain (exon 18–28, amino acids 669–1210). The cytoplasmic domain includes the tyrosine kinase (exons 18–24, amino acids 669–979) and C-terminal tail (exon 24–28, amino acids 980–1210) domains
[35].
To date, several EGFR variants have been studied in humans
[36][37][38][36,37,38]. In addition to the full-length isoform (isoform A), cells can also produce alternate EGFR isoforms that lack the intracellular domain
[39]. The different roles of these alternative isoforms have not yet been established, but some have now been shown to play crucial roles
[40]. Therefore,
thwe
researchers aanalyzed the different EGFR transcript variants reported in the genomic and protein databases for zebrafish and humans, as summarized in and .
Table 1. List of zebrafish EGFR isoforms in the Ensembl database. ID, transcript name, transcript length, protein name and protein length for all zebrafish EGFR isoforms listed in the Ensembl database.
|
Ensembl ID
|
Transcript
|
Length (nt)
|
Protein (Uniprot)
|
Length (aa)
|
Table 2. List of human EGFR isoforms in the Ensembl and NCBI genomic databases. ID, transcript name, transcript length, protein name and protein length for all human EGFR isoforms listed in the Ensembl (top) and NCBI (bottom) databases. Isoforms highlighted in green are common to both databases.
|
Ensembl ID
|
Transcript
|
Length (nt)
|
Protein (Uniprot)
|
Length (aa)
|
|
egfra-201
|
ENSDART00000108964.5
|
1868
|
F1RBY7
|
503
|
|
egfra-202 |
|
EGFR-201
|
ENST00000275493.7
|
9905
|
P0053-1
|
1210
|
|
|
ENSDART00000128514.2
|
1881
|
F1RA48
|
|
EGFR-202
|
ENST00000342916.7
|
2239
|
P00533-4
|
389
|
|
| 628 |
|
egfra-203
|
|
|
EGFR-203
| ENSDART00000136906.3
|
ENST00000344576.6
2437
|
|
2864
F1R671
|
760
|
|
| P00533-3 |
|
| 705
|
egfra-205
|
ENSDART00000147261.3
|
3168
|
|
EGFR-204
|
ENST00000420316.6
|
| F1QU74
|
243
|
|
| 1570 |
|
P00533-2
|
405
|
egfra-206
|
ENSDART00000150499.3
|
|
EGFR-205
|
ENST00000450046.1
|
3007
|
|
691
F1Q7X2
|
C9JYS6
625
|
|
|
| 128 |
|
egfra-207
|
|
EGFR-206
|
ENSDART00000164152.3
|
6151
|
A0A0R4IFV9
|
1191
|
ENST00000454757.6 |
|
|
|
|
5464 |
|
|
|
| E9PFD7 |
|
| 1165
|
|
EGFR-207
|
ENST00000455089.5
|
3844
|
Q504U8
|
1091
|
|
NCBI ID
|
Transcript
|
Length (nt)
|
Protein (NCBI)
|
Length (aa)
|
|
Isoform A
|
NM_005228.5
|
9905
|
NP_005219.1
|
1210
|
|
Isoform G
|
NM_001346899.2
|
9770
|
NP_001346899.2
|
1165
|
|
Isoform I
|
NM_001346941.2
|
9104
|
NP_001333870.1
|
943
|
|
Isoform F
|
NM_001346898.2
|
3983
|
NP_001333827.1
|
1136
|
|
Isoform E
|
NM_001346897.2
|
3848
|
NP_001333826.1
|
1091
|
|
Isoform D
|
NM_201284.2
|
2872
|
NP_958441.1
|
705
|
|
Isoform B
|
NM_201282.2
|
2254
|
NP_958439.1
|
628
|
|
Isoform C
|
NM_201283.2
|
1575
|
NP_958440.1
|
405
|
|
Isoform H
|
NM_001346900.2
|
9676
|
NP_001333829.1
|
1157
|
Analysis of zebrafish genomic data version 11 on the Ensembl website, RGCz11;
www.ensembl.org (accessed on 4 March 2021), revealed identifying six transcripts for egfra (egfra-201; 202; 203; 205; 206 and 207) (), and one transcript, which retained an intron and, therefore, is not listed in . Teleosts have experienced a fish-specific genome duplication
[41], and, therefore, around one-third of the zebrafish genes are duplicated. In the zebrafish, a putative paralog named egfrb has been identified, but to date, no isoforms or transcript variants have been reported
[42], and only one paper has reported the expression of the egfrb paralog in zebrafish
[43]. The latest human genomic data on the Ensembl website (
www.ensembl.org, accessed on 4 March 2021), human genome version (GRCh38.p13), revealed identifying 7 reported transcripts (EGFR-201 to 207) ranging from 1210 amino acids in length to 128 amino acids in length (). Ensembl human genomic data analysis also revealed the identification of two processed transcripts (561 and 452 bp, respectively) and one retained intron (665 bp), none of which are translated into a protein and, therefore, were excluded from .
NCBI analysis of the latest human EGFR transcript sequences (updated 7 March 2021) within the National Library of Medicine protein database revealed the existence of 9 EGFR transcripts encoding proteins from 1210 amino acids in length to 405 amino acids in length (). Comparison of the Ensembl and NCBI data sets revealed that 7 transcripts are common in both databases (highlighted in green in ), while the reported transcript EGFR-205 is unique to the Ensembl database, and isoforms I, F and H are only present in the NCBI database ().