Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 2 by Sirius Huang and Version 1 by Ivana Škrlec.
The growing emergence of antimicrobial resistance represents a global problem that not only influences healthcare systems but also has grave implications for political and economic processes. As the discovery of novel antimicrobial agents is lagging, one of the solutions is innovative therapeutic options that would expand our armamentarium against this hazard. Compounds of interest in many such studies are antimicrobial peptides (AMPs), which actually represent the host’s first line of defense against pathogens and are involved in innate immunity. They have a broad range of antimicrobial activity against Gram-negative and Gram-positive bacteria, fungi, and viruses, with specific mechanisms of action utilized by different AMPs.
- antimicrobial peptides
- antimicrobial resistance
- antimicrobial effects
- antibacterial activity
1. Introduction
Antimicrobial peptides (AMPs) have played a key role in numerous scientific studies intending to find new antimicrobial agents and active substances [1]. The need for new antimicrobial drugs is increasingly recognized, driven by the growing global burden of antimicrobial resistance and a significant increase in infections that cannot be treated with existing antibiotics [2,3,4][2][3][4]. A particular problem is the ESCAPE group of pathogens, which includes Escherichia coli, Staphylococcus aureus, Klebsiella pneumoniae, Streptococcus pneumoniae, Acinetobacter baumannii, and Pseudomonas aeruginosa, which, according to data from 2019, are responsible for more than 250,000 antimicrobial-related deaths resistance [5]. Antimicrobial resistance is associated with high mortality rates. In addition, it significantly impacts the effectiveness of antimicrobial agents as it reduces the efficacy of treatment and prolongs the time of infection in patients [5]. Additionally, another group of pathogens associated with antimicrobial resistance is responsible for another 100,000 to 250,000 deaths, consisting of M. tuberculosis, Enterococcus faecium, Enterobacters spp., Streptococcus agalactiae, S. typhi, and Enterococcus faecalis. All this influenced the increase in treatment costs due to the use of more expensive therapies [5]. It is important to note that a significantly higher degree of resistance is present in Gram-negative bacteria. Accordingly, treating infections caused by this group of pathogens is more complicated. Namely, many first-line antibiotics, such as vancomycin, rifampicin, and others that are successful in treating infections caused by Gram-positive pathogens due to their ineffectiveness against Gram-negative pathogens, are not used in the treatment of infections caused by this group of pathogens. The problem is the permeability of the lipopolysaccharide (LPS) of the outer membrane and porins because they largely limit the entry of antibiotics into the cell [6]. Gram-negative opportunistic pathogens could be resistant to commercial antibiotics due to the LPS of the outer membrane [7]. Cationic AMPs provide new possibilities in treating infections caused by Gram-negative pathogens because they can often permeabilize the anionic LPS of the outer membrane, which leads to cell lysis and opens the way to action on Gram-negative bacteria. Likewise, AMPs can act as enhancers of the antimicrobial activity of conventional antibiotics on Gram-negative pathogens [8]. However, some Gram-negative bacteria have an innate resistance to cationic AMPs [9] associated with changes in the LPS of the outer membrane [7,9][7][9]. AMPs are known to be widespread [10]; they are involved in innate immunity and represent the first line of defense against pathogens in the host [11]. They show a wide range of antimicrobial activity against Gram-negative and Gram-positive bacteria, fungi (particularly yeasts), and viruses [12,13][12][13]. Significantly, they have a low tendency to induce drug resistance, thus providing us with the opportunity to use them as a specific class of antibiotics [14,15][14][15]. However, we have to be cognizant that the organism’s protection from pathogens depends on how they distribute in tissues and whether external administration is free from toxicity issues [16].
2. Antimicrobial Effects of AMPs
2.1. Antibacterial Activity of AMPs
As the primary goal in researching AMPs was the fight against (increasingly resistant) bacteria, some scientists emphasize this need and call them antibacterial peptides (ABP) [34,35][17][18]. They can destroy bacteria by breaking the cell wall and membrane of the bacteria, intracellular action, through a combination of dual destruction mechanisms, and by acting on the bacterial biofilm [36,37][19][20]. By binding to lipid II, which is part of the peptidoglycan molecule and an essential factor in cell wall synthesis, ABPs prevent cell wall formation. In addition to controlling its assembly, it can destroy the already-formed cell wall [37,38,39,40][20][21][22][23]. Acting on the membrane is possible in several ways. The barrel-shaped and toroidal pore models can be defined as transmembrane pore models. Vertical insertion of the peptide into the membrane while promoting peptide–peptide lateral interaction is typical of the barrel-shaped pore model [37,41,42][20][24][25]. In the toroidal pore model, peptides are also vertically embedded in the membrane. However, here the pores are created by both peptide and lipid groups. The most crucial difference between these two models is that the barrel model’s hydrophobic and hydrophilic bilayers remained intact [37,42,43][20][25][26]. The model that does not create pores through the membrane and in which specific peptide–peptide interactions are not necessary belongs to the so-called carpet model. It is characterized by adherence of ABP along the bacterial membrane and its adsorption until a particular concentration is reached. After crossing the tolerance threshold, a detergent-like model appears, and a loss of integrity and opening of the bacterial membrane happens [13,37,43,44,45][13][20][26][27][28]. Many peptides work by destroying intracellular functions, and in that way, they kill bacteria. Some important ones inhibit DNA, RNA, and protein synthesis [42,46,47,48][25][29][30][31]. The development of research and the discovery of peptides with antimicrobial effects show that specific peptides do not have only one impact model (Figure 1). On the contrary, most of them share a combination of mechanisms [37,42,43,49,50][20][25][26][32][33].
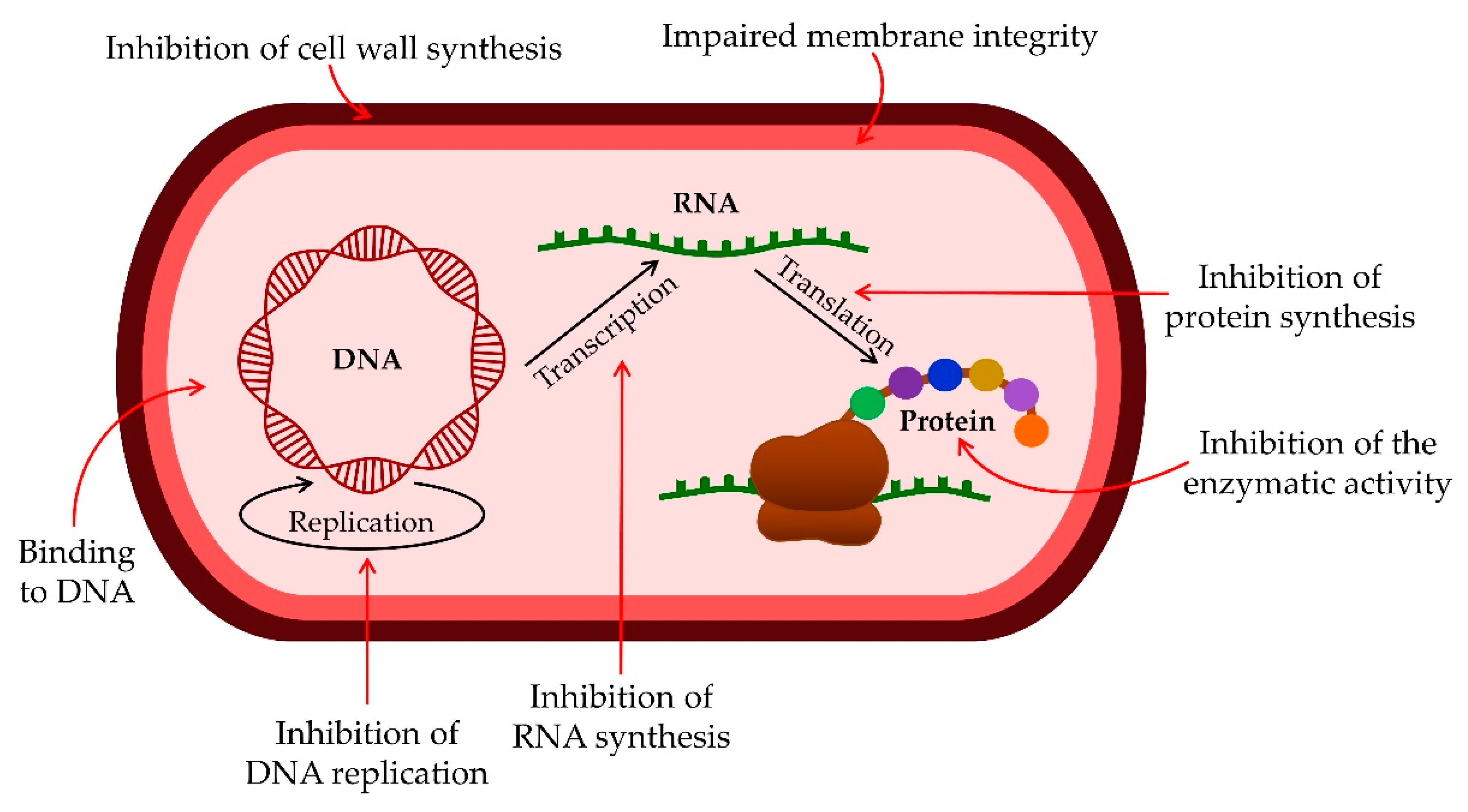
Figure 1.
Antibacterial activity mechanisms of AMPs.
AMPs can act on biofilm at all stages of its development. They can inhibit biofilm formation by disrupting the signaling pathway of bacteria cells. By encouraging bacteria to produce guanosine tetraphosphate (ppGpp) and pentaphosphate (pppGpp) with limiting nutrients in the biofilm, the synthesis of nucleic acids is inhibited. Another effect is a reduction in the expression of binding protein transport genes necessary for forming a bacterial biofilm. Additionally, ABP can destroy the already-formed biofilm by acting on the membrane potential of bacteria [37,43,51,52][20][26][34][35].
2.2. Antiviral Activity of AMPs
Due to the increasing resistance of viruses and the limited effect of common drugs, antiviral peptides play an essential role as therapeutic agents [43][26]. AMPs with antiviral activity are referred to as antiviral peptides (AVPs) [53,54][36][37]. There are several different mechanisms by which antiviral peptides block viruses at various stages of their cycle [34][17]. Some AVPs can destabilize the viral membrane or prevent infection by neutralizing the virus by integrating it into the viral envelope and cell membrane [2,55][2][38]. Similarly, they can bind to viral glycoproteins, after which the viruses can no longer bind to the host cell surface [2[2][39],56], while some bind to host cell receptors, preventing the virus from binding to its target receptor and thus inactivating it (e.g., heparan for herpes simplex virus). Some block viral attachment and prevent membrane fusion in the virus cell (e.g., influenza virus) [55,57][38][40]. An essential mechanism of action of AVP on the influenza virus is the regulation of the human immune system with the increased expression of cytokines and chemokines on the antigen complex, activation of cells of the immune system, and inactivation of viral pathogens (e.g., H1N1 virus) [58,59,60][41][42][43]. In addition, AVP blocks its activity by acting on the replication cycle [61][44]. AVP shows similar effects to those described for influenza virus in inhibiting human immunodeficiency virus 1 (HIV-1), herpes simplex virus 1 (HSV-1), herpes simplex virus 2 (HSV-2), hepatitis B virus (HBV), and hepatitis C virus (HCV) infections [61][44]. The target site of AVPs can be DNA and RNA [62[45][46],63], and the goal of their action is to destroy the viral envelope, leading to its instability, as in the Junin virus (JV), HIV-1, and HSV-2 [64][47]. Another way to inhibit viruses is to modify or interfere with cell signaling pathways [65][48], such as GF-17 (17-mer-derived peptide from human cathelicidin LL-37) and BMAP-18 (bovine myeloid antimicrobial peptide-18), which can inactivate the Zika virus (ZIKV) by interfering with interferon type 1 signaling [54][37]. Recently, there has been an increasing number of studies on the antiviral effect of AVPs on coronaviruses (such as SARS-CoV). In short, the current dogma of this type of research endeavors is based on AVP’s inhibitory effect on the viral cell membrane and its inability to enter the host cell [66,67,68,69][49][50][51][52].
2.3. Antifungal Activity of AMPs
The excessive and improper use of systemic antibiotics, immunosuppressive therapy, chemotherapy, and radiotherapy have increased fungal infections in the general population [70,71,72][53][54][55]. The limited choice of antifungal therapy, which includes only four chemical types of systemic antifungals—polyenes, azoles, echinocandins, and flucytosines [71,73][54][56]—as well as the increase in the resistance of specific strains to the mentioned antifungal drugs, leads to increased morbidity and mortality, but also a dire need for alternative solutions in antifungal treatment [70,71,74][53][54][57].
Antifungal peptides (AFPs) could be a promising therapy for fungal infections [71,75][54][58]. Most AFPs achieve their activity through membrane-associated mechanisms and specific targets. Differences in fungal membranes are sphingolipid composition, PI content, and ergosterol as the main membrane sterol. Specific targets such as glucosylceramides, mannosyldiinositol phosphorylceramide, or a fungal protein target enable high selectivity and avoid resistance to therapy [76][59].
AFPs can be classified in accordance with a number of criteria, such as structure, mode of action, or origin [77][60]. However, the most accepted classification is according to the origin, which divides peptides into natural, semisynthetic, and synthetic ones [78][61].
Natural AFPs are produced by different species like Archaea, Bacteria, and Eukaryotes [77][60]. Natural AFPs have an α-helix structure, β-hairpin or sheet, or combination of α-helix and β-sheet, and depending on the amino acid in the most significant composition. They are classified as glycine-rich, arginine-rich, proline-rich, histidine-rich, and tryptophan-rich peptides [79][62].
Semisynthetic and synthetic peptides are designed to improve pharmacological properties and reduce immunogenicity and side effects caused by natural peptides [77][60]. Biophysical characteristics such as net charge, stereospecificity, hydrophobicity, secondary structure, peptide length, and amphipathicity determine the antifungal activity of peptides [80][63]. For example, an increase in the positive net charge can cause a stronger action on the membrane. Likewise, increased hydrophobicity and amphipathicity lead to increased antifungal activity [77][60].
The most abundant peptides harmful to the fungal biofilm are peptides of mammalian origin, defensins, cathelicidins, and histatins [71][54]. Defensins are isolated from not only mammals but also plants [81][64]. They are structurally organized as α-helix and triple-stranded antiparallel β-sheet, which are connected by disulfide bonds that ensure the stability of the structure, even in extreme conditions [82][65]. Defensins from vertebrate animals are cationic and amphipathic peptides and can be divided into two subfamilies, α-defensins and β-defensins [83][66]. Human α-defensin 6 (HD6) prevents the adhesion of Candida albicans (C. albicans) to human intestinal epithelial cells and, thus, biofilm formation [84][67]. β-defensin-1 displays inhibitory activity against germinating conidia of Aspergillus fumigatus [85][68]. Synthetic defensin-like peptides like α-defensin-3, β-defensin-1, β-defensin-3, and PG-1 express antifungal activity against Cryptococcus neoformans biofilms, including both planktonic cells and mature biofilm [86][69]. Cathelicidins are cationic peptides isolated from different species of mammals, consisting of 12–80 amino acids [71,87][54][70]. The human antimicrobial peptide LL-37 was proven to inhibit C. albicans cell adhesion on polystyrene and silicon surfaces, and BMAP-28, a bovine antimicrobial peptide, was able to reduce the number of C. albicans adherent cells on silicone surfaces and inhibit its mature biofilm [88,89,90][71][72][73]. Histatins are human salivary peptides, first isolated from human parotid saliva, with polar and hydrophilic properties and α-helix structural conformation in organic solutions [91][74]. Histatin-5 (Hst-5) was proven to inhibit biofilm formation of C. albicans on acrylic dentures in vitro [92][75] and, in another study, inhibit biofilm formation and planktonic cells of C. albicans and Candida glabrata on methyl methacrylate disks [93][76].
2.4. Immunomodulatory Activity of AMPs
AMPs play a significant role in immunomodulation and inflammation control [94,95][77][78] (Figure 2). The mechanisms of action of AMPs in immune modulation involve various immune responses [96][79]. The three main families of AMPs in humans are defensins, histatins, and cathelicidins. Based on the arrangement of disulfide bonds, defensins are divided into α-defensins and β-defensins, and they are produced by lymphocytes, neutrophils, and epithelial cells of mucous membranes and skin [24][80].
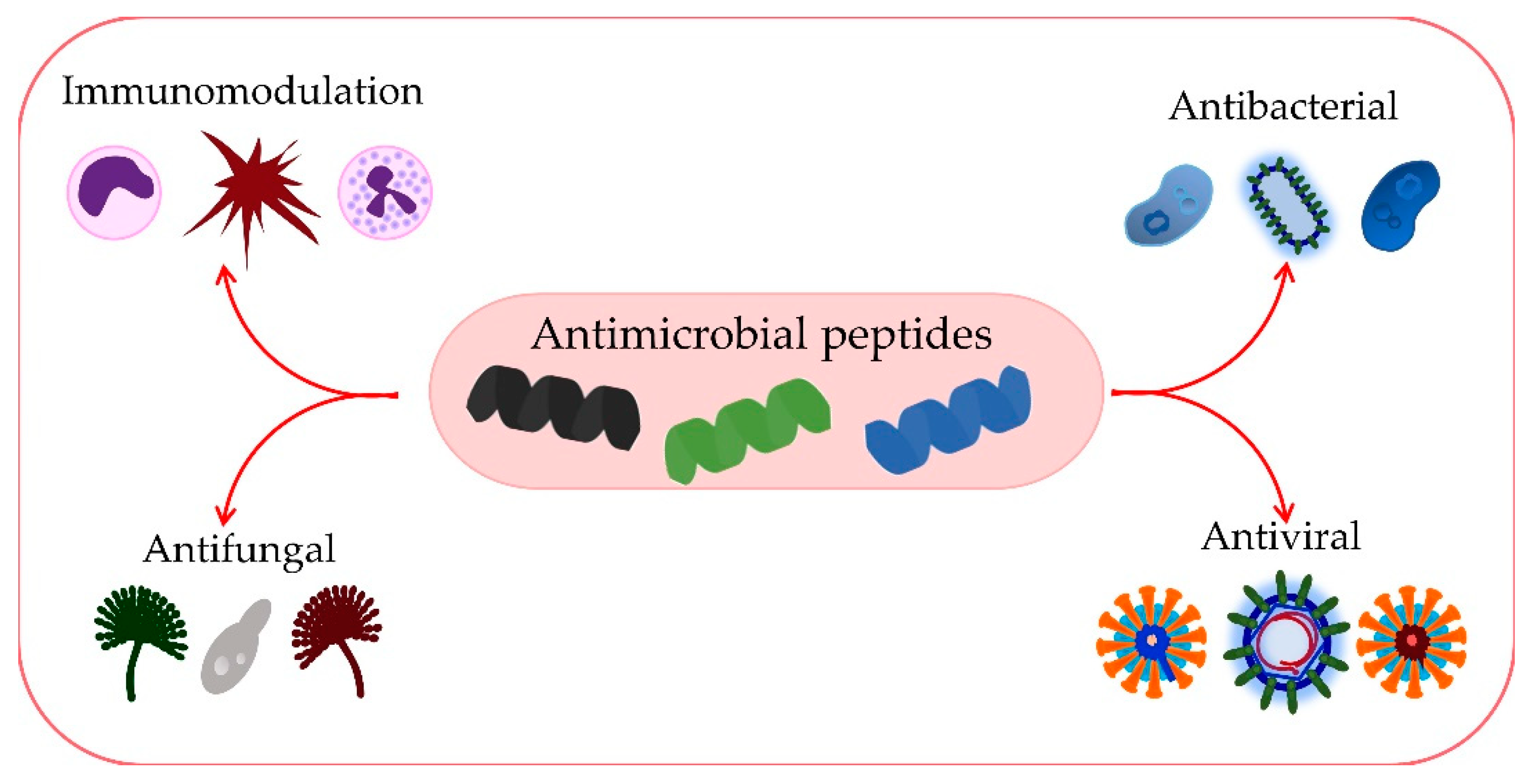
Figure 2.
Numerous functions of AMPs.
2.4.1. Defenses
Since human α-defensins 1-4, most commonly expressed by neutrophils, are called neutrophil peptides 1-4 (HNP) [97][81], together with lysozyme, proteases, and other proteins, they participate in the destruction of bacterial pathogens. This gives them exceptional importance in the immune system [98][82]. Human α-defensins 5 and 6 are produced and secreted mainly by Paneth cells located at the base of Lieberkühn’s crypts in the small intestine and by epithelial cells of the male and female genital organs [99][83]. In addition, some human AMPs (β-defensins, LL-37) can attract immune cells such as white blood cells, dendritic cells, and mast cells [100][84]. Beta defensins originate from the epithelial cells of the skin and mucosa of mammals [101][85]. Thus far, six types of human β-defensins have been isolated and designated as hBD 1-6 [102][86].
Regarding antimicrobial activity and expression level, hBD-1, hBD-2, and hBD-3 defensins were characterized [101][85]. Epithelial cells constitutively express hBD-1, hBD-2, and hBD-3, but after stimulation by microorganisms and pro-inflammatory cytokines [103][87]. The role of β-defensin in connecting adaptive and innate immunity is significant [104][88]. More specifically, hBD-3 has the ability to rapidly enter TLR4-stimulated macrophages and dampen the expression of pro-inflammatory genes [105][89]. The role of β-defensin peptides is multifunctional, so in addition to the defensive, antimicrobial function, they also have a clearly expressed immunomodulatory function [106][90]. Since they are ubiquitous across mucosal surfaces, they are considered essential factors in homeostasis and health [107][91].
The administration of human peptides has been shown to have multiple protective effects in an in vivo model of infection [19][92]. Furthermore, some AMPs can act as regulatory molecules, as evidenced by in vitro research showing that defensins can attract phagocytes and lymphocytes to the site of infection, induce fibroblast proliferation, and modify ion flow in epithelial cells [108][93]. In addition, AMPs can induce bacterial lysis, promote macrophage phagocytosis, prevent the spread of infection, induce mitosis of fibroblasts and epithelial cells, and promote fibroblast growth to improve wound healing [1]. Finally, based on the ability of AMPs to stimulate complement activation and the production of cytokines and antibodies, the influence of AMPs on humoral immunity is currently being further evaluated [98][82].
2.4.2. Histatins
AMPs, histatins, contain large amounts of histidine amino acids [17][94]. They were isolated from human parotid salivary glands [109][95] and are characterized by antifungal and antimicrobial properties [110][96]. Human saliva contains Histatin-1, and Histatin-3, which are derived from the available genes HTN1 and HTN3. They differ in the number of amino acids and molecular weight [111][97]. Histatin-5 is derived from histatin-3 and contains an N-terminal for which it is highly reactive and has an affinity for binding with metals [112][98]. They are characterized by antifungal and antimicrobial properties, while the effects of histatin on immune system cells are unknown [113][99].
2.4.3. Cathelicidins
Cathelicidins are constitutively expressed at low levels in epithelial cells, skin, and mucosal surfaces and are released to a considerable extent in response to infections, especially by granulocytes and mononuclear phagocytes [114][100]. Cathelicidins in in vitro conditions have an apparent antimicrobial effect on parasites, fungi, bacteria, and enveloped viruses [115][101]. Cationic cathelicidins have different mechanisms of action on the cell, damaging the integrity of the negatively charged membrane, which results in cell death [116][102]. They can act on intracellular processes by promoting protein breakdown, weakening enzymes’ role, and affecting RNA and DNA synthesis [117][103]. Numerous studies have observed that in the presence of glycosaminoglycans, salt, bacterial DNA, and mucin, i.e., under physiological conditions, the action of LL-37 is inhibited even at high peptide concentrations [88,118][71][104]. Based on this, it can be concluded that, in vivo conditions, most cathelicidins probably do not have a direct bactericidal effect. However, they still have great importance for preventing microbial infections due to their immunomodulatory effect [98,100][82][84]. The ability of cathelicidins to attract many adaptive and innate immune cells to inflammatory sites by modulating the expression of chemokines and chemokine receptors should undoubtedly be highlighted [119][105]. Thus, cathelicidins can act on various immune cells as direct chemoattractants. This effect is enhanced by their indirect impact on inducing the upregulation of chemokines and chemokine receptors on leukocytes [114,119,120][100][105][106].
References
- Lei, J.; Sun, L.C.; Huang, S.; Zhu, C.; Li, P.; He, J.; Mackey, V.; Coy, D.H.; He, Q.Y. The antimicrobial peptides and their potential clinical applications. Am. J. Transl. Res. 2019, 11, 3919–3931.
- Talapko, J.; Škrlec, I. The Principles, Mechanisms, and Benefits of Unconventional Agents in the Treatment of Biofilm Infection. Pharmaceuticals 2020, 13, 299.
- Burnham, J.P. Climate change and antibiotic resistance: A deadly combination. Ther. Adv. Infect. Dis. 2021, 8, 2049936121991374.
- Giacomini, E.; Perrone, V.; Alessandrini, D.; Paoli, D.; Nappi, C.; Esposti, L.D. Evidence of Antibiotic Resistance from Population-Based Studies: A Narrative Review. Infect. Drug Resist. 2021, 14, 849–858.
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Robles Aguilar, G.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655.
- Bhattacharjya, S.; Mohid, S.A.; Bhunia, A. Atomic-Resolution Structures and Mode of Action of Clinically Relevant Antimicrobial Peptides. Int. J. Mol. Sci. 2022, 23, 4558.
- Shaka, M.; Arias-Rojas, A.; Hrdina, A.; Frahm, D.; Iatsenko, I. Lipopolysaccharide-mediated resistance to host antimicrobial peptides and hemocyte-derived reactive-oxygen species are the major Providencia alcalifaciens virulence factors in Drosophila melanogaster. PLoS Pathog. 2022, 18, e1010825.
- Bhattacharjya, S.; Straus, S.K. Design, Engineering and Discovery of Novel α-Helical and β-Boomerang Antimicrobial Peptides against Drug Resistant Bacteria. Int. J. Mol. Sci. 2020, 21, 5773.
- Ghimire, J.; Guha, S.; Nelson, B.J.; Morici, L.A.; Wimley, W.C. The Remarkable Innate Resistance of Burkholderia bacteria to Cationic Antimicrobial Peptides: Insights into the Mechanism of AMP Resistance. J. Membr. Biol. 2022; in press.
- Gan, B.H.; Gaynord, J.; Rowe, S.M.; Deingruber, T.; Spring, D.R. The multifaceted nature of antimicrobial peptides: Current synthetic chemistry approaches and future directions. Chem. Soc. Rev. 2021, 50, 7820–7880.
- Marshall, J.S.; Warrington, R.; Watson, W.; Kim, H.L. An introduction to immunology and immunopathology. Allergy Asthma. Clin. Immunol. 2018, 14, 49.
- Reddy, K.V.R.; Yedery, R.D.; Aranha, C. Antimicrobial peptides: Premises and promises. Int. J. Antimicrob. Agents 2004, 24, 536–547.
- Huan, Y.; Kong, Q.; Mou, H.; Yi, H. Antimicrobial Peptides: Classification, Design, Application and Research Progress in Multiple Fields. Front. Microbiol. 2020, 11, 582779.
- León-Buitimea, A.; Garza-Cárdenas, C.R.; Garza-Cervantes, J.A.; Lerma-Escalera, J.A.; Morones-Ramírez, J.R. The Demand for New Antibiotics: Antimicrobial Peptides, Nanoparticles, and Combinatorial Therapies as Future Strategies in Antibacterial Agent Design. Front. Microbiol. 2020, 11, 1669.
- Larsson, D.G.J.; Flach, C.F. Antibiotic resistance in the environment. Nat. Rev. Microbiol. 2022, 20, 257–269.
- Pollard, A.J.; Bijker, E.M. A guide to vaccinology: From basic principles to new developments. Nat. Rev. Immunol. 2021, 21, 83–100.
- Moravej, H.; Moravej, Z.; Yazdanparast, M.; Heiat, M.; Mirhosseini, A.; Moosazadeh Moghaddam, M.; Mirnejad, R. Antimicrobial Peptides: Features, Action, and Their Resistance Mechanisms in Bacteria. Microb. Drug Resist. 2018, 24, 747–767.
- Boman, H.G. Antibacterial peptides: Basic facts and emerging concepts. J. Intern. Med. 2003, 254, 197–215.
- Fjell, C.D.; Hiss, J.A.; Hancock, R.E.W.; Schneider, G. Designing antimicrobial peptides: Form follows function. Nat. Rev. Drug Discov. 2011, 11, 37–51.
- Luo, Y.; Song, Y. Mechanism of Antimicrobial Peptides: Antimicrobial, Anti-Inflammatory and Antibiofilm Activities. Int. J. Mol. Sci. 2021, 22, 11401.
- Koch, A.L. Bacterial wall as target for attack: Past, present, and future research. Clin. Microbiol. Rev. 2003, 16, 673–687.
- Münch, D.; Sahl, H.G. Structural variations of the cell wall precursor lipid II in Gram-positive bacteria—Impact on binding and efficacy of antimicrobial peptides. Biochim. Biophys. Acta 2015, 1848, 3062–3071.
- Wenzel, M.; Chiriac, A.I.; Otto, A.; Zweytick, D.; May, C.; Schumacher, C.; Gust, R.; Albada, H.B.; Penkova, M.; Krämer, U.; et al. Small cationic antimicrobial peptides delocalize peripheral membrane proteins. Proc. Natl. Acad. Sci. USA 2014, 111, E1409–E1418.
- Barreto-Santamaría, A.; Curtidor, H.; Arévalo-Pinzón, G.; Herrera, C.; Suárez, D.; Pérez, W.H.; Patarroyo, M.E. A New Synthetic Peptide Having Two Target of Antibacterial Action in E. coli ML35. Front. Microbiol. 2016, 7, 2006.
- Haney, E.F.; Mansour, S.C.; Hancock, R.E.W. Antimicrobial Peptides: An Introduction. Methods Mol. Biol. 2017, 1548, 3–22.
- Di Somma, A.; Moretta, A.; Canè, C.; Cirillo, A.; Duilio, A. Antimicrobial and Antibiofilm Peptides. Biomolecules 2020, 10, 652.
- Yeaman, M.R.; Yount, N.Y. Mechanisms of antimicrobial peptide action and resistance. Pharmacol. Rev. 2003, 55, 27–55.
- Ciumac, D.; Gong, H.; Hu, X.; Lu, J.R. Membrane targeting cationic antimicrobial peptides. J. Colloid Interface Sci. 2019, 537, 163–185.
- Milletti, F. Cell-penetrating peptides: Classes, origin, and current landscape. Drug Discov. Today 2012, 17, 850–860.
- Brogden, K.A. Antimicrobial peptides: Pore formers or metabolic inhibitors in bacteria? Nat. Rev. Microbiol. 2005, 3, 238–250.
- Neundorf, I. Antimicrobial and Cell-Penetrating Peptides: How to Understand Two Distinct Functions Despite Similar Physicochemical Properties. Adv. Exp. Med. Biol. 2019, 1117, 93–109.
- Yan, J.; Wang, K.; Dang, W.; Chen, R.; Xie, J.; Zhang, B.; Song, J.; Wang, R. Two hits are better than one: Membrane-active and DNA binding-related double-action mechanism of NK-18, a novel antimicrobial peptide derived from mammalian NK-lysin. Antimicrob. Agents Chemother. 2013, 57, 220–228.
- Ko, S.J.; Kang, N.H.; Kim, M.K.; Park, J.; Park, E.; Park, G.H.; Kang, T.W.; Na, D.E.; Park, J.B.; Yi, Y.E.; et al. Antibacterial and anti-biofilm activity, and mechanism of action of pleurocidin against drug resistant Staphylococcus aureus. Microb. Pathog. 2019, 127, 70–78.
- Wolz, C.; Geiger, T.; Goerke, C. The synthesis and function of the alarmone (p)ppGpp in firmicutes. Int. J. Med. Microbiol. 2010, 300, 142–147.
- Otto, M. Bacterial evasion of antimicrobial peptides by biofilm formation. Curr. Top. Microbiol. Immunol. 2006, 306, 251–258.
- Maiti, B.K. Potential Role of Peptide-Based Antiviral Therapy against SARS-CoV-2 Infection. ACS Pharmacol. Transl. Sci. 2020, 3, 783–785.
- Li, X.; Zuo, S.; Wang, B.; Zhang, K.; Wang, Y. Antimicrobial Mechanisms and Clinical Application Prospects of Antimicrobial Peptides. Molecules 2022, 27, 2675.
- Ahmed, A.; Siman-Tov, G.; Hall, G.; Bhalla, N.; Narayanan, A. Human Antimicrobial Peptides as Therapeutics for Viral Infections. Viruses 2019, 11, 704.
- Vilas Boas, L.C.P.; Campos, M.L.; Berlanda, R.L.A.; de Carvalho Neves, N.; Franco, O.L. Antiviral peptides as promising therapeutic drugs. Cell. Mol. Life Sci. 2019, 76, 3525–3542.
- Skalickova, S.; Heger, Z.; Krejcova, L.; Pekarik, V.; Bastl, K.; Janda, J.; Kostolansky, F.; Vareckova, E.; Zitka, O.; Adam, V.; et al. Perspective of Use of Antiviral Peptides against Influenza Virus. Viruses 2015, 7, 5428–5442.
- Lee, H.; Lee, Y.; Kim, J.; An, J.; Lee, S.; Kong, H.; Song, Y.; Lee, C.K.; Kim, K. Modulation of the gut microbiota by metformin improves metabolic profiles in aged obese mice. Gut Microbes 2018, 9, 155–165.
- Barlow, P.G.; Svoboda, P.; Mackellar, A.; Nash, A.A.; York, I.A.; Pohl, J.; Davidson, D.J.; Donis, R.O. Antiviral activity and increased host defense against influenza infection elicited by the human cathelicidin LL-37. PLoS ONE 2011, 6, e25333.
- Holani, R.; Babbar, A.; Blyth, G.A.D.; Lopes, F.; Jijon, H.; McKay, D.M.; Hollenberg, M.D.; Cobo, E.R. Cathelicidin-mediated lipopolysaccharide signaling via intracellular TLR4 in colonic epithelial cells evokes CXCL8 production. Gut Microbes 2020, 12, 1785802.
- Hoffmann, J.; Schneider, C.; Heinbockel, L.; Brandenburg, K.; Reimer, R.; Gabriel, G. A new class of synthetic anti-lipopolysaccharide peptides inhibits influenza A virus replication by blocking cellular attachment. Antiviral Res. 2014, 104, 23–33.
- Horne, W.S.; Wiethoff, C.M.; Cui, C.; Wilcoxen, K.M.; Amorin, M.; Ghadiri, M.R.; Nemerow, G.R. Antiviral cyclic D,L-alpha-peptides: Targeting a general biochemical pathway in virus infections. Bioorg. Med. Chem. 2005, 13, 5145–5153.
- Mulder, K.C.L.; Lima, L.A.; Miranda, V.J.; Dias, S.C.; Franco, O.L. Current scenario of peptide-based drugs: The key roles of cationic antitumor and antiviral peptides. Front. Microbiol. 2013, 4, 321.
- Albiol Matanic, V.C.; Castilla, V. Antiviral activity of antimicrobial cationic peptides against Junin virus and herpes simplex virus. Int. J. Antimicrob. Agents 2004, 23, 382–389.
- He, M.; Zhang, H.; Li, Y.; Wang, G.; Tang, B.; Zhao, J.; Huang, Y.; Zheng, J. Cathelicidin-derived antimicrobial peptides inhibit Zika virus through direct inactivation and interferon pathway. Front. Immunol. 2018, 9, 722.
- Bakovic, A.; Risner, K.; Bhalla, N.; Alem, F.; Chang, T.L.; Weston, W.; Harness, J.A.; Narayanan, A. Brilacidin Demonstrates Inhibition of SARS-CoV-2 in Cell Culture. Viruses 2021, 13, 271.
- Bhattacharya, R.; Gupta, A.M.; Mitra, S.; Mandal, S.; Biswas, S.R. A natural food preservative peptide nisin can interact with the SARS-CoV-2 spike protein receptor human ACE2. Virology 2021, 552, 107–111.
- Liscano, Y.; Oñate-Garzón, J.; Ocampo-Ibáñez, I.D. In Silico Discovery of Antimicrobial Peptides as an Alternative to Control SARS-CoV-2. Molecules 2020, 25, 5535.
- Zhang, R.; Jiang, X.; Qiao, J.; Wang, Z.; Tong, A.; Yang, J.; Yang, S.; Yang, L. Antimicrobial peptide DP7 with potential activity against SARS coronavirus infections. Signal Transduct. Target. Ther. 2021, 6, 140.
- De Cesare, G.B.; Cristy, S.A.; Garsin, D.A.; Lorenz, M.C. Antimicrobial peptides: A new frontier in antifungal therapy. mBio 2020, 11, e02123-20.
- Oshiro, K.G.N.; Rodrigues, G.; Monges, B.E.D.; Cardoso, M.H.; Franco, O.L. Bioactive Peptides against Fungal Biofilms. Front. Microbiol. 2019, 10, 2169.
- Vallabhaneni, S.; Chiller, T.M. Fungal Infections and New Biologic Therapies. Curr. Rheumatol. Rep. 2016, 18, 29.
- Chowdhary, A.; Sharma, C.; Meis, J.F. Azole-resistant aspergillosis: Epidemiology, molecular mechanisms, and treatment. J. Infect. Dis. 2017, 216, S436–S444.
- Fisher, M.C.; Hawkins, N.J.; Sanglard, D.; Gurr, S.J. Health and Food Security—TCLocal. Science 2018, 742, 739–742.
- di Luca, M.; Maccari, G.; Nifosí, R. Treatment of microbial biofilms in the post-antibiotic era: Prophylactic and therapeutic use of antimicrobial peptides and their design by bioinformatics tools. Pathog. Dis. 2014, 70, 257–270.
- Rautenbach, M.; Troskie, A.M.; Vosloo, J.A. Antifungal peptides: To be or not to be membrane active. Biochimie 2016, 130, 132–145.
- Fernández de Ullivarri, M.; Arbulu, S.; Garcia-Gutierrez, E.; Cotter, P.D. Antifungal Peptides as Therapeutic Agents. Front. Cell. Infect. Microbiol. 2020, 10, 105.
- Lucca, A.J. De Expert Opinion on Investigational Drugs Antifungal peptides: Potential candidates for the treatment of fungal infections. Expert Opin. Investig. Drugs 2000, 9, 273–299.
- Bondaryk, M.; Staniszewska, M.; Zielińska, P.; Urbańczyk-Lipkowska, Z. Natural Antimicrobial Peptides as Inspiration for Design of a New Generation Antifungal Compounds. J. Fungi 2017, 3, 46.
- Akkam, Y. A review of antifungal peptides: Basis to new era of antifungal drugs. Jordan J. Pharm. Sci. 2016, 9, 51–75.
- Cools, T.L.; Struyfs, C.; Cammue, B.P.; Thevissen, K. Antifungal plant defensins: Increased insight in their mode of action as a basis for their use to combat fungal infections. Future Microbiol. 2017, 12, 441–454.
- Shafee, T.M.A.; Lay, F.T.; Hulett, M.D.; Anderson, M.A. The Defensins Consist of Two Independent, Convergent Protein Superfamilies. Mol. Biol. Evol. 2016, 33, 2345–2356.
- Parisi, K.; Shafee, T.M.A.; Quimbar, P.; van der Weerden, N.L.; Bleackley, M.R.; Anderson, M.A. The evolution, function and mechanisms of action for plant defensins. Semin. Cell Dev. Biol. 2019, 88, 107–118.
- Chairatana, P.; Chiang, I.L.; Nolan, E.M. Human α-Defensin 6 Self-Assembly Prevents Adhesion and Suppresses Virulence Traits of Candida albicans. Biochemistry 2017, 56, 1033–1041.
- Ballard, E.; Yucel, R.; Melchers, W.J.G.; Brown, A.J.P.; Verweij, P.E.; Warris, A. Antifungal activity of antimicrobial peptides and proteins against Aspergillus fumigatus. J. Fungi 2020, 6, 65.
- Martinez, L.R.; Casadevall, A. Cryptococcus neoformans cells in biofilms are less susceptible than planktonic cells to antimicrobial molecules produced by the innate immune system. Infect. Immun. 2006, 74, 6118–6123.
- Zanetti, M.; Gennaro, R.; Romeo, D. Cathelicidins: A novel protein family with a common proregion and a variable C-terminal antimicrobial domain. FEBS Lett. 1995, 374, 1–5.
- Ridyard, K.E.; Overhage, J. The potential of human peptide ll-37 as an antimicrobial and anti-biofilm agent. Antibiotics 2021, 10, 650.
- Risso, A.; Braidot, E.; Sordano, M.C.; Vianello, A.; Macrì, F.; Skerlavaj, B.; Zanetti, M.; Gennaro, R.; Bernardi, P. BMAP-28, an Antibiotic Peptide of Innate Immunity, Induces Cell Death through Opening of the Mitochondrial Permeability Transition Pore. Mol. Cell. Biol. 2002, 22, 1926–1935.
- Scarsini, M.; Tomasinsig, L.; Arzese, A.; D’Este, F.; Oro, D.; Skerlavaj, B. Antifungal activity of cathelicidin peptides against planktonic and biofilm cultures of Candida species isolated from vaginal infections. Peptides 2015, 71, 211–221.
- Oppenheim, F.G.; Xu, T.; McMillian, F.M.; Levitz, S.M.; Diamond, R.D.; Offner, G.D.; Troxler, R.F. Histatins, a novel family of histidine-rich proteins in human parotid secretion. Isolation, characterization, primary structure, and fungistatic effects on Candida albicans. J. Biol. Chem. 1988, 263, 7472–7477.
- Pusateria, C.R.; Monacoa, E.A.; Edgertona, M. Sensitivity of Candida albicans Biofilm Cells Grown on Denture Acrylic to Antifungal Proteins and Chlorhexidine. Arch. Oral Biol. 2009, 54, 588–594.
- Konopka, K.; Dorocka-Bobkowska, B.; Gebremedhin, S.; Düzgüneş, N. Susceptibility of Candida biofilms to histatin 5 and fluconazole. Antonie van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2010, 97, 413–417.
- van der Does, A.M.; Hiemstra, P.S.; Mookherjee, N. Antimicrobial Host Defence Peptides: Immunomodulatory Functions and Translational Prospects. Adv. Exp. Med. Biol. 2019, 1117, 149–171.
- Kang, H.K.; Lee, H.H.; Seo, C.H.; Park, Y. Antimicrobial and Immunomodulatory Properties and Applications of Marine-Derived Proteins and Peptides. Mar. Drugs 2019, 17, 350.
- Kumar, P.; Kizhakkedathu, J.N.; Straus, S.K. Antimicrobial Peptides: Diversity, Mechanism of Action and Strategies to Improve the Activity and Biocompatibility In Vivo. Biomolecules 2018, 8, 4.
- Zhang, Q.Y.; Yan, Z.B.; Meng, Y.M.; Hong, X.Y.; Shao, G.; Ma, J.J.; Cheng, X.R.; Liu, J.; Kang, J.; Fu, C.Y. Antimicrobial peptides: Mechanism of action, activity and clinical potential. Mil. Med. Res. 2021, 8, 48.
- Ehmann, D.; Koeninger, L.; Wendler, J.; Malek, N.P.; Stange, E.F.; Wehkamp, J.; Jensen, B.A.H. Fragmentation of Human Neutrophil α-Defensin 4 to Combat Multidrug Resistant Bacteria. Front. Microbiol. 2020, 11, 1147.
- Guryanova, S.V.; Ovchinnikova, T.V. Immunomodulatory and Allergenic Properties of Antimicrobial Peptides. Int. J. Mol. Sci. 2022, 23, 2499.
- Ouellette, A.J. Paneth cell α-defensins in enteric innate immunity. Cell. Mol. Life Sci. 2011, 68, 2215–2229.
- Pahar, B.; Madonna, S.; Das, A.; Albanesi, C.; Girolomoni, G. Immunomodulatory Role of the Antimicrobial LL-37 Peptide in Autoimmune Diseases and Viral Infections. Vaccines 2020, 8, 517.
- Meade, K.G.; O’Farrelly, C. β-Defensins: Farming the Microbiome for Homeostasis and Health. Front. Immunol. 2019, 9, 2072.
- Semple, F.; Dorin, J.R. β-Defensins: Multifunctional modulators of infection, inflammation and more? J. Innate Immun. 2012, 4, 337–348.
- Ghosh, S.K.; Feng, Z.; Fujioka, H.; Lux, R.; McCormick, T.S.; Weinberg, A. Conceptual Perspectives: Bacterial Antimicrobial Peptide Induction as a Novel Strategy for Symbiosis with the Human Host. Front. Microbiol. 2018, 9, 302.
- Machado, L.R.; Ottolini, B. An evolutionary history of defensins: A role for copy number variation in maximizing host innate and adaptive immune responses. Front. Immunol. 2015, 6, 115.
- Candela, M.E.; Allsop, D.J.P.; Carter, R.N.; Semple, F.; Kilanowski, F.; Webb, S.; Taggart, D.; Mullan, H.J.; McHugh, B.J.; Dockrell, D.H.; et al. Classical macrophage polarisation is limited by human β-defensin-3 via an autocrine IL-4 dependent process. bioRxiv 2021. Preprint.
- Xu, D.; Lu, W. Defensins: A Double-Edged Sword in Host Immunity. Front. Immunol. 2020, 11, 764.
- Zheng, D.; Liwinski, T.; Elinav, E. Interaction between microbiota and immunity in health and disease. Cell Res. 2020, 30, 492–506.
- Moretta, A.; Scieuzo, C.; Petrone, A.M.; Salvia, R.; Manniello, M.D.; Franco, A.; Lucchetti, D.; Vassallo, A.; Vogel, H.; Sgambato, A.; et al. Antimicrobial Peptides: A New Hope in Biomedical and Pharmaceutical Fields. Front. Cell. Infect. Microbiol. 2021, 11, 668632.
- Gera, S.; Kankuri, E.; Kogermann, K. Antimicrobial peptides—Unleashing their therapeutic potential using nanotechnology. Pharmacol. Ther. 2021, 232, 107990.
- Lachowicz, J.I.; Szczepski, K.; Scano, A.; Casu, C.; Fais, S.; Orrù, G.; Pisano, B.; Piras, M.; Jaremko, M. The Best Peptidomimetic Strategies to Undercover Antibacterial Peptides. Int. J. Mol. Sci. 2020, 21, 7349.
- Komatsu, T.; Watanabe, K.; Hamada, N.; Helmerhorst, E.; Oppenheim, F.; Lee, M.C. Il Association between Antimicrobial Peptide Histatin 5 Levels and Prevalence of Candida in Saliva of Patients with Down Syndrome. Antibiotics 2021, 10, 494.
- Sharma, P.; Chaudhary, M.; Khanna, G.; Rishi, P.; Kaur, I.P. Envisaging Antifungal Potential of Histatin 5: A Physiological Salivary Peptide. J. Fungi 2021, 7, 1070.
- Bastos, P.; Trindade, F.; da Costa, J.; Ferreira, R.; Vitorino, R. Human Antimicrobial Peptides in Bodily Fluids: Current Knowledge and Therapeutic Perspectives in the Postantibiotic Era. Med. Res. Rev. 2018, 38, 101–146.
- Norris, H.L.; Kumar, R.; Ong, C.Y.; Xu, D.; Edgerton, M. Zinc Binding by Histatin 5 Promotes Fungicidal Membrane Disruption in C. albicans and C. glabrata. J. Fungi 2020, 6, 124.
- Lee, S.M.; Son, K.N.; Shah, D.; Ali, M.; Balasubramaniam, A.; Shukla, D.; Aakalu, V.K. Histatin-1 Attenuates LPS-Induced Inflammatory Signaling in RAW264.7 Macrophages. Int. J. Mol. Sci. 2021, 22, 7856.
- van Harten, R.M.; van Woudenbergh, E.; van Dijk, A.; Haagsman, H.P. Cathelicidins: Immunomodulatory Antimicrobials. Vaccines 2018, 6, 63.
- Zhang, L.; Wu, W.K.K.; Gallo, R.L.; Fang, E.F.; Hu, W.; Ling, T.K.W.; Shen, J.; Chan, R.L.Y.; Lu, L.; Luo, X.M.; et al. Critical Role of Antimicrobial Peptide Cathelicidin for Controlling Helicobacter pylori Survival and Infection. J. Immunol. 2016, 196, 1799–1809.
- Benfield, A.H.; Henriques, S.T. Mode-of-Action of Antimicrobial Peptides: Membrane Disruption vs. Intracellular Mechanisms. Front. Med. Technol. 2020, 2, 610997.
- Lin, L.; Chi, J.; Yan, Y.; Luo, R.; Feng, X.; Zheng, Y.; Xian, D.; Li, X.; Quan, G.; Liu, D.; et al. Membrane-disruptive peptides/peptidomimetics-based therapeutics: Promising systems to combat bacteria and cancer in the drug-resistant era. Acta Pharm. Sin. B 2021, 11, 2609–2644.
- Barańska-Rybak, W.; Sonesson, A.; Nowicki, R.; Schmidtchen, A. Glycosaminoglycans inhibit the antibacterial activity of LL-37 in biological fluids. J. Antimicrob. Chemother. 2006, 57, 260–265.
- Alford, M.A.; Baquir, B.; Santana, F.L.; Haney, E.F.; Hancock, R.E.W. Cathelicidin Host Defense Peptides and Inflammatory Signaling: Striking a Balance. Front. Microbiol. 2020, 11, 1902.
- Choi, K.Y.G.; Mookherjee, N. Multiple immune-modulatory functions of cathelicidin host defense peptides. Front. Immunol. 2012, 3, 149.
More