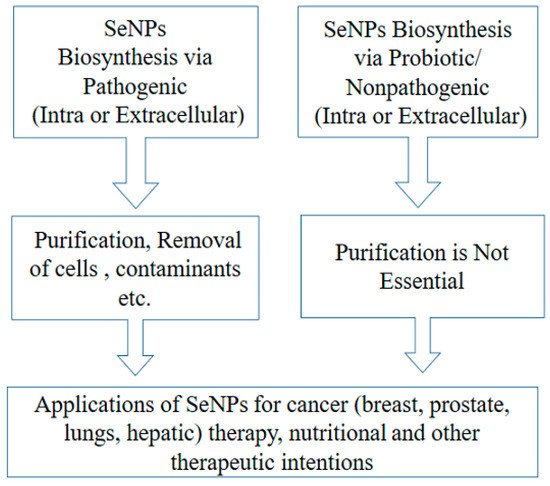
1. Presumed Anticancer Mechanism
The anticancer effects of SeNPs remain unclear to date; however, the presumed differential effects and cellular anticancer mechanism of SeNPs are outlined (
Figure 1,
Figure 2 and
Figure 3). In general, biogenic SeNPs have shown more differential cytotoxicity on cancer cells compared with normal cells
[1][2][107,117]. The chemopreventive influence of SeNPs and their presumed anticancer mechanisms were recently well evaluated by Menon et al.
[3][108] and Khurana et al.
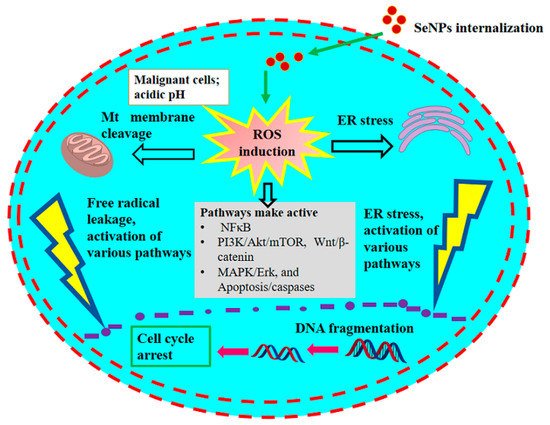
[1][107]. Cancer cells have an acidic pH with redox perturbations, and this internal environment leads to a preoxidative transformation of SeNPs, triggering an increased production of free radicals. This leads on the one hand to a disruption of the mitochondrial membrane, causing mitochondrial (Mt) proteins to leak out and on the other hand to a stress of the endoplasmic reticulum (ER). Disruption of the Mt membrane results in the efflux of various proteins and triggers apoptosis through the activation of caspases (a family of protease enzymes that play an essential role in programmed cell death). This stress state of the cell coordinates the activation of several molecular signaling pathways, including MAPK/Erk, PI3K/Akt/mTOR, NFκB, Wnt/β-catenin, and apoptosis pathways. The NFκB pathway triggers oxidative stress and disrupts cellular homeostasis through inflammation. The PI3K/Akt/mTOR, MAPK/Erk, VEGF, and Wnt/β-catenin signaling pathways are important for oncogenic signaling, and their modulation by SeNPs impairs cellular proliferation and hinders growth-promoting signaling in the cancer cell microenvironment. Moreover, SeNPs reduced angiogenic signaling in tumor cells, further inhibiting proliferation and growth. The consolidation of these disruptive cellular events begins with the destruction of DNA, leading to cell cycle arrest and eventually cell death.
Figure 1. Process handling of biogenic SeNPs and their application as anticancer agents.
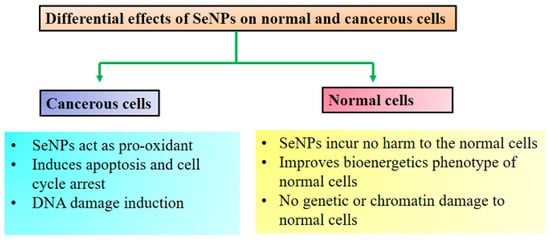
Figure 2. Differential anticancer effect of SeNPs. Adopted from Khurana et al. [4]. Differential anticancer effect of SeNPs. Adopted from Khurana et al. [106].
Figure 3. The anticancer mechanism of SeNPs in cancerous cells. Updated as from Khurana et al. [4]. The anticancer mechanism of SeNPs in cancerous cells. Updated as from Khurana et al. [106].
2. Effective against Breast Cancer
Breast carcinoma is the most common malignancy in women throughout the world, with significant regional and racial disparities
[5][6][7][128,129,130]. The incidence of this disease in Pakistan is 2.5 times higher than in India and Iran. Approximately 1 million cases of breast cancer in women are reported annually, mainly in industrialized countries
[8][131]. In the past decades, it has become clear that matrix metalloproteinases (MMPs) play an important role in tumor development, metastasis, and incidence. MMPs can serve as good biomarkers for breast carcinoma, and their levels are related to the stage of the disease
[9][10][132,133]. Proteolysis of the basement membrane and extracellular matrix, particularly collagen IV, is one of the essential processes involved in breast cancer metastasis
[11][134], and this proteolytic degradation occurs through the action of various MMPs. In addition to metastasis, MMPs are also involved in tumor growth, angiogenesis, and vasculogenesis
[9][132].
Bacillus sp. MSh-1-produced SeNPs with a particle size of 80–220 nm showed cytotoxic and inhibitory effects on human MMP-2 expression in HT-1080 (fibrosarcoma cell line) in a dose-dependent manner
[12][33]. This inhibitory effect of SeNPs on the expression of MMPs may also be helpful in other carcinomas, as overexpression of MMP2 has been similarly found in bladder cancer
[13][135], oral carcinoma
[14][136], CRC
[15][16][137,138], lung cancer
[17][139], prostate cancer
[18][140], as well as gastric cancer
[19][141]. Se nanoparticles (80–220 nm) generated with the same species (
Bacillus sp. MSh-1) were found to be almost 6-fold less cytotoxic to breast cancer cell MCF-7 compared to SeO
2 at concentrations of 6.7 µg/mL. The same SeNPs also showed better DPPH scavenging activity and lower performance at 200 µg/mL, making them the expected choice for breast melanoma
[20][31].
SeNPs produced by
Acinetobacter sp. SW30 showed a selective cytotoxic effect on breast cancer cells (4T1, MCF-7) but not on non-cancer cells (NIH/3T3, HEK293). Therefore, they were recommended as a good choice for breast cancer cell selection. Oral administration (2.5 × 10
8 CFU/mL) of
Lactobacillus plantarum (ATCC 8014)-enriched SeNPs to mice demonstrated an effective immune response by promoting pro-inflammatory cytokines TNF-α, IFN-γ, and IL-2 in splenocytes and enhanced NK activity (natural killing) of cells
[21][22][119,120]. Survival was significantly improved and tumor volume decreased compared to 4T1 breast cancer-bearing control mice
[23][54]. Similarly, the purified SeNPs produced by the same probiotic bacteria were orally administered to 4T1 breast cancer-bearing mice to study the immune response
[24][55]. In this
res
earchtudy, the production of Th1 cytokines such as IL-12 and IFN-γ in spleen cells was enhanced in the test mice administered SeNPs. In addition, delayed hypersensitivity reaction (DTH) and survival were also significantly increased compared to control.
Yazdi et al.
[25][53] used
Lactobacillus brevis to produce SeNP-enriched biomass. A total of 0.5 mL orally administered biomass induced an efficient immune response by increasing IL-17 and IFN-γ levels with a remarkable increase in DTH responses and NK cytotoxicity in tumor-induced BALB/c mice. In addition, they also increased the survival rate and decreased the metastasis of liver tumors. Therefore, SeNPs containing biomass have been suggested as suitable candidates for the upcoming prevention and immunotherapy of breast tumors.
Another study conducted by Yazdi et al. showed that injection of purified SeNPs (produced by the same species;
Lactobacillus brevis) and 4T1 cell antigen vaccine improved immune responses by increasing serum IL-2, IL-12, and IFN-γ levels and reduced TGF-β
[26][56]. In addition, the vaccine demonstrated a stronger DTH response, reduced tumor volume, and prolonged survival in the mouse breast cancer model
[27][57].
Similarly, Yazdi et al. investigated the immune responses with purified SeNPs produced by
Lactobacillus brevis after oral administration of 100 µg/day in BALB/c mice after inducing breast cancer
[27][57]. It was shown that cellular immunomodulatory components such as granzyme B, IL-2, IL-12, and IFN-γ were significantly improved in the mice treated with both SeNPs and crude antigens of 4T1 cells compared with the other groups (
p < 0.05). However, TGF-β levels decreased in certain mice
[27][57].
In the same year, Faghfuri et al. also studied SeNPs (50–250 nm) derived from
Lactobacillus brevis [28][58]. Their results also showed an increase in an immunomodulatory component such as IFN-γ and the IFN-γ/IL-4 ratio at all doses administered compared to control doses. In addition, a lower tumor volume and prolonged survival were observed at a higher dose (200 mg/day) of SeNPs.
3. Effective against Prostate Cancer
Although prostate cancer is usually a slowly developing carcinoma, it is the third leading cause of cancer deaths in men, even in developed countries
[29][142]. Each form of Se has been attributed to an anticarcinogenic effect with different mechanisms of action, and most of them have been restrained in prostate cancer
[30][31][143,144]. SeNPs (40–180 nm) prepared by Sonkusre and Cameotra
[32][36] using
Bacillus licheniformis have been investigated for their potential use against prostate cancer. The harvested SeNPs induced necrosis in LNCaP-FGC cells without affecting red blood cell integrity at a nominal concentration of 2 μg/mL. A real-time gene expression study showed overexpression of TNF (tumor necrotic factor) and interferon regulatory factor (IRF) and a decrease in androgen receptor (AR) and prostate-specific antigen (PSA) expression. Significantly lower toxicity was observed after oral administration of a ten-fold higher concentration (50 mg Se/kg) of the same SeNPs in C3H/HeJ mice compared to 5 mg Se/kg L-selenomethionine
[33][121].
Nanoparticles (40–180 nm) produced by
Bacillus licheniformis JS2 promoted late necrosis or apoptosis in PC3 at a concentration of 2 µg/mL SeNPs, but no marked necrosis or apoptosis was detected in hPBMCs (normal cells) at the same concentration
[34][122]. Later, similar SeNPs were reproduced by Sonkusre
[22][120], and the anticancer mechanism was further investigated
[32][36]. The results showed that the comparable SeNPs promoted ROS-mediated necroptosis in PC-3 cells at a Se concentration of 2 μg/mL by cellular internalization. The qPCR study showed an increase in gene expression of necroptosis-related IRF1 and TNF. Improved expression of RIP1 protein was also detected at the translational level. In addition, cell viability was significantly improved in the presence of the necroptosis inhibitor necrostatin-1.
4. Effective against Lung Cancer
Lung cancer is considered one of the most recurrent malignancies worldwide
[35][145]. Treatment outcomes are among the poorest of all cancers, and survival is only 10–20% in five years
[36][146]. SeNPs (50–80 nm particle size) produced anaerobically by
Bacillus licheniformis ATCC 10716 have been shown to have a chemopreventive effect on lung cancer. The
res
earchtudy aimed to test SeNPs as a chemotherapeutic agent against lung cancer under the influence of iron (III) nitrilotriacetate in male Wistar rats. Pretreatment (0.2 mg Se/kg body weight) with SeNPs significantly restored glutathione levels, catalase, and superoxide dismutase (SOD) activities and improved oxidative damage parameters such as lipid peroxidation, nitric oxide, and inflammatory factors such as C-reactive protein and TNF-α levels with improvement in the histological analysis of lung tissue and retraction of hyperplasia cells
[37][52].
Bao et al. synthesized SeNPs (100–500 nm) using
Bacillus oryziterrae and investigated their anticancer activity
[34][122]. The results showed considerable inhibition of lung cancer cell line H157 in a dose-dependent manner, supporting a potential application of SeNPs in lung cancer treatment.
5. Effective against Hepatic Carcinoma
Hepatocellular carcinoma (HCC) is a cause of numerous cancer-related deaths worldwide. It is considered the most common carcinoma of the liver, originating from hepatocytes and occurring in nearly 80% of liver cancer cases
[38][147]. Biogenic SeNPs with a particle size of 50–80 nm produced anaerobically by the probiotic
Lactobacillus casei ATCC 393 showed significant inhibition of tumor development in the human liver cell line HepG2 and ameliorated diquat-induced oxidative stress in IPEC-J2 cells
[39][124]. The same method for producing Se nanoparticles was applied and additionally enriched with proteins and polysaccharides to further investigate their antioxidant and hepatoprotective effects. The results of the
res
earchtudy showed that SeNPs produced by
L. casei 393 promoted apoptosis of HepG2 cells via caspase signaling cascade and endocytosis of SeNPs. Moreover, at a concentration of <25 μg Se/mL, they showed no cytotoxic effect on NCM460 proliferation and growth. Similarly, oxidative damage induced by H
2O
2 or diquat was ameliorated in intestinal epithelial cells, malondialdehyde (MDA) level was decreased, and GPx (glutathione peroxidase) activity was increased in the culture medium. These results indicate that the capped SeNPs produced by the probiotic
L. casei 393 have a substantial effect on HCC
[40][125].
6. Effective against Colon Cancer
Colorectal cancer (CRC) is the third most common disease in men and the second most common cancer in women. More than 1 million new cases are diagnosed each year, and nearly 0.5 million people die from it annually
[41][148]. Even in the United States, 106,180 new cases of colon cancer and 44,850 cases of rectal cancer were reported in the current year, while 52,580 deaths were due to colon and rectal cancer combined
[42][149]. SeNPs (170 –550 nm) produced by the probiotic strain
Lactobacillus casei ATCC 393 were shown to inhibit colon cancer cell growth in vitro and in vivo. The specific SeNPs showed antiproliferative activity against CT26 colon cancer in mice. They also tended to induce apoptosis with an increased level of ROS in colon cancer cells, HT29
[43][118]. The same research group also confirmed the colorectal anticancer activity of SeNPs of the same species. These biogenic SeNPs have pro-apoptotic activity and the ability to promote immunogenic cell death (ICD) of colon cancer cells. In this in vitro study, the biomarkers involved in ICD (translocation of calreticulin and ERp57, the release of HMGB1 and ATP) were revealed, and the secretion of pro-inflammatory cytokines from cells treated with SeNPs was detected
[21][119]. In addition, a study by Xu et al.
[39][124] showed that SeNPs (50–80 nm) synthesized from the same
Lactobacillus strain had a protective effect on the colon by stimulating the growth/proliferation of normal human IPEC-J2 cells, colon epithelial cells (NCM460), and human acute monocytic leukemia cell (THP-1)-derived macrophagocytes.
7. Effective as Antioxidants
All mammalian selenoproteins contain Se in the form of the amino acid selenocysteine (Sec), which is encoded by the UGA triplet. There are two forms of tRNA[Ser]Sec, which are essential for the synthesis of all selenoproteins. The tRNA[Ser]Sec isoforms are both the site of Sec synthesis and the adaptor molecules, which recognize the appropriate UGA codons in selenoprotein mRNAs. Twenty-two known eukaryotic selenoproteins are organized into distinct selenoprotein groups on the basis of the location and functional properties of Sec. GSH-Px, selenoprotein P, type I iodothyronine 5′-deiodinase (DI-I), and thioredoxin reductase (TR) have been characterized in animals and humans. Approximately half of the characterized selenoproteins have been implicated in having antioxidant functions. Thus, increased risks of human diseases associated with Se deficiency may be attributable to increased oxidative stress. Moreover, this oxidative stress and the resulting oxidative damage are important contributors to the formation and progression of cancer. However, Se supplementation can increase the level of enzymatic proteins, prevent the accumulation of free radical species, and reduce cellular damage
[44][45][46][47][48][49][50][150,151,152,153,154,155,156]. ROS and RNS (reactive nitrogen species) are free radicals that form naturally and play an important role in normal cell physiology. However, in higher concentrations, these radicals can be harmful and damage key cellular components, especially proteins, DNA, and cell membranes
[51][52][53][157,158,159]. The damage caused by reactive free radicals, especially DNA damage, is the cause of cancer and other health disorders
[54][55][160,161]. Se is usually referred to as a nutritional antioxidant, but this effect is mainly attributed to selenium-containing proteins and not to pure Se
[56][162]. SeNPs have been shown to have better antioxidant activity than other chemical forms of Se and also reduce the possibility of its toxicity
[57][40].
Exopolysaccharide-capped biogenic SeNPs accumulated directly by
Bacillus paralicheniformis SR14 (294 nm average particle size) showed greater antioxidant effects in scavenging ABTS, DPPH, and superoxide free radicals, but not OH radicals. In vitro studies with IPEC-J2 also showed remarkable cytoprotection of the same SeNPs against H
2O
2-induced oxidative stress, as they suppressed ROS formation. These effects suggest major antioxidant and cytoprotective properties of SeNPs for normal cells
[58][126].
In another study by Xu et al., SeNPs were suggested to be a promising Se supplement with antioxidant and anti-inflammatory properties
[58][126]. In this study, SeNPs (38–152 nm particle size) were biosynthesized using the probiotic
Lactococcus lactis NZ9000 in an economical and environmentally friendly manner. These SeNPs significantly improved MDA (malondialdehyde) concentration and decreased GPx and total SOD activity in IPEC-J2 exposed to H
2O
2. Similarly, they prevented the H
2O
2-induced decrease in transepithelial electrical resistance and increased the FITC-dextran flux of IPEC-J2. Moreover, SeNPs decreased the increase in ROS, decreased mitochondrial membrane potential and ATP, and supported intestinal epithelial permeability in H2O2-exposed IPEC-J2 cells. Moreover, pretreated SeNPs attenuated the cytotoxic effect of
E. coli (ETEC) K88 on IPEC-J2 cells and preserved the integrity of the intestinal epithelial barrier by upregulating occludin and claudin-1 expression along with a reduction in inflammatory cytokines.
Qiao et al. investigated in vivo the antioxidant parameters of biogenic SeNPs (50–80 nm) produced by the probiotic Lactobacillus casei ATCC 393 on diquat-induced intestinal barrier disruption and nuclear mechanisms in C57BL/6 mice
[59]. Their results showed that oral administration of SeNPs significantly suppressed the increase in serum levels of alanine aminotransferase (ALT), aspartate aminotransferase (AST), diamine oxidase (DAO), and D-lactic acid promoted by diquat and improved the overall activities of SOD, GPx, and thioredoxin reductase (TrxR) in the jejunum and serum. In addition, they improved the number of goblet cells, decreased the formation of ROS, maintained mitochondrial function, and improved the expression of claudin-1 and occludin in the jejunum compared with the induced oxidative stress group model. In addition, SeNPs also stimulated nuclear factor Nrf2 and increased NADPH dehydrogenase and heme oxygenase. These results suggest that the above-mentioned SeNPs produced by
L. casei ATCC 393 can protect intestinal barrier functions by counteracting oxidative damage induced by Nrf2 signaling.