1. Introduction
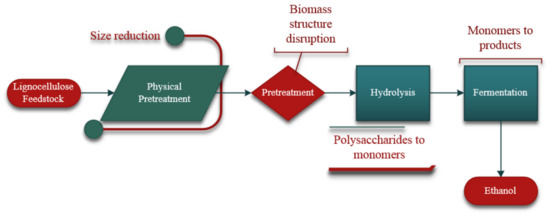
A representative diagram of ethanol production from lignocellulosic feedstock is shown in Figure 1. The process includes mechanical size reduction, followed by pretreatment in which the biomass structure is disrupted and separated further. Hydrolysis, which is the focus of this work, occurs next. Polysaccharides are broken down into monomers such as glucose, xylose, and arabinose. During fermentation, simple sugars are converted to ethanol.
Figure 1.
Biorefinery process of ethanol production from lignocellulosic feedstock.
2. Pretreatment
Pretreatment is the step in which lignocellulosic biomass is converted from being recalcitrant to a form ready for enzymatic hydrolysis
[1][23]. An efficient pretreatment process must be able to recover complete lignocellulosic components and produce less degradation byproducts. In addition, it must be feasible and effective on various biomass quantities and types
[1][23]. The different types of pretreatment methods used with lignocellulosic biomass can be classified into physical, chemical, and biological pretreatments.
2.1. Physical Pretreatment
Most biochemical conversion processes use physical pretreatment as a first step. This step serves to reduce the particle size, which increases the surface area to volume ratio and reduces the degree of polymerization and crystallinity of the biomass, thus enhancing the conversion rate
[2][24]. The enzymatic hydrolysis of cotton cellulose was enhanced to produce glucose yields of up to 99.8% over 50 h when the particle size was reduced from 25 to 0.78 µm
[3][25]. The several types of physical treatments, such as chipping, shredding, milling, and grinding, produce different final particle sizes. In general, decreasing the particle size below 0.3 mm has been proven to increase the glucose conversion yield
[4][26].
2.2. Chemical Pretreatment
Acidic Pretreatment
Hemicellulose and cellulose are partially solubilized by both dilute and concentrated acids. However, concentrated acids are not favored due to their severe effects on the biomass and the process. Their effects include cellulose degradation and inhibitor production
[5][27]. Dilute acids are used within a concentration range of 0.5% to 2.5% and a temperature range of 100 to 200 °C
[5][27]. Various studies have investigated the enhancement of the enzymatic hydrolysis step via dilute sulfuric acid pretreatment
[6][7][28,29]. Pretreating rice straw with 1% (
w/
w) sulfuric acid enhanced the efficiency of enzymatic hydrolysis to 70% by increasing the biomass pore volume. This resulted in a combined glucose and xylose yield of 83% after 72 h
[8][30]. Although enzymatic hydrolysis is enhanced by this type of pretreatment, the partial hydrolysis of cellulose results in more crystalline cellulose. This produces lower conversion yields than other pretreatment methods
[9][31].
Alkaline Pretreatment
In contrast to acid pretreatment, the alkaline reagent used in alkaline pretreatment interacts with, breaks down, and isolates only lignin from the biomass during alkaline pretreatment. This can provide a simple, inexpensive way of enhancing enzymatic hydrolysis
[10][32]. The use of alkaline compounds, such as sodium hydroxide, sodium carbonate, and ammonia, makes this pretreatment method superior to other methods because these compounds are non-corrosive chemicals that require mild conditions
[10][32]. In addition, this method is highly selective for lignin removal and retains cellulose and hemicellulose intact. The absence of inhibitor production enhances the fermentation step. It was found that pretreating rice straw with ultrasound-assisted alkaline (NaOH) improved the digestible cellulose yield by a factor of 3.5 compared to untreated biomass
[11][33]. The suggested pretreatment method was found to increase the surface area accessible for cellulase and to increase the porosity compared to the same biomass treated with heat only. However, this pretreatment method generally increases the crystallinity index. This is due primarily to lignin removal rather than to structural changes within the cellulose
[10][32]. However, the degree of polymerization was found to decrease when pretreatment was performed using a NaOH concentration below 8 wt%. Such low concentrations caused separation in the cellulose lattice
[12][34]. In addition, pretreating sugarcane bagasse with 20% aqueous ammonia for 48 h at 50 °C was found to result in 57.3% total sugar release during the subsequent enzymatic hydrolysis
[13][35].
Oxidative Pretreatment
Oxidizing agents such as oxygen, ozone, and hydrogen peroxide are used to remove lignin. However, partial breakdown of some of the hemicellulose is observed. In addition, the aforementioned agents are not selective for lignin. They can attack cellulose and lead to the production of byproducts such as aliphatic aldehydes and aliphatic organic acids, which inhibit the subsequent enzymatic hydrolysis
[14][36]. This pretreatment method can be combined with other methods to improve the degradation of lignocellulose and enhance enzymatic hydrolysis. For example, corn stover was pretreated in a two-stage process; the first stage was pretreatment with dilute hydrochloric acid (1 wt%) for 40 min at 120 °C. This was followed by alkaline wet oxidative pretreatment with 12.6 wt% ammonium hydroxide under pressurized oxygen (3 MPa) at 130 °C for the same duration. This two-stage process was found to remove approximately 86% of lignin and produce xylan and glucan yields of 82.8% and 71.5%, respectively
[15][37].
2.3. Physicochemical Pretreatment
This class of pretreatment changes the structure of the biomass both physically and chemically. Several types, such as solvent fractionation, steam explosion, liquid hot water, and carbon dioxide explosion, are used.
Solvent Fractionation
Solvent fractionation is the partial solubilization of lignocellulosic components by breaking down hydrogen bonds between fibrils. This occurs because different lignocellulosic components have different solubilities in different solvents
[2][24]. This type of pretreatment involves the use of organic solvents, ionic liquids, or phosphoric acid.
Organic solvents, such as ethanol in the presence of an acid catalyst, are used to extract lignin from the biomass and thus reduce the crystallinity
[16][38]. However, some properties of these solvents restrict their application as a pretreatment method. For example, the use of organic solvents with low boiling points, such as ethanol, acetone, methanol, and ethyl acetate
[16][38], necessitates operating at high pressure. In addition, safety issues should be considered when using flammable solvents
[17][39]. However, the fractionation of corn stover biomass using ethanol resulted in a 91% glucan content after lignin removal, while the fractionation of giant miscanthus and wheat straw with ethanol was not efficient
[18][40]. In addition, the use of organic amines, such as polyamine, as ethanol fractionation catalysts has been shown to aid in boosting the delignification of corn stover biomass. This produced a lignin removal of 82% and a sugar yield of 83%
[19][41]. Furthermore, combining sulfuric acid and ethanol to perform wheat straw pretreatment was found to enhance the extraction of fermentable sugars to 89%, which is better than the other organic solvents tested, such as methanol, butanol, acetone, and diethylene glycol
[20][42]. Although different solvents have been reported for lignocellulose fractionation, their residence time, biomass loading, byproduct production, and structural disruption are all factors to consider when selecting the most convenient pretreatment method. For example, using a cellulose solvent such as concentrated phosphoric acid produced better structural disruption and a 97% glucan yield in 24 h. This can be compared to dilute sulfuric acid, which achieved 84% in 72 h. However, the effect of using such concentrated acids on inhibitor production was not reported
[21][22][43,44].
Ionic liquids (ILs) such as 1-allyl-3-methylimidazolium chloride [AMIMCl], -allyl-3-methylimidazolium acetate [EMIM][AC], and 1-butyl-3-methylimidazolium chloride [BMIMCl] have been shown to solubilize cellulose from biomass effectively
[23][45]. Due to the presence of anions, such as chloride, cellulose can bind with the ionic liquid via hydrogen. Cellulose can then be recovered using an antisolvent, such as water, which breaks down these bonds and enables recovery of the used IL
[23][45]. Thus, pretreatment with ILs is an area of research interest
[24][46]. Sugar yields of 89% and 87% from sugarcane bagasse and wheat straw, respectively, were achieved when both biomasses were pretreated with [EMIM][AC]
[25][47]. Pretreatment of wheat straw with 1-ethyl-3-methyl-imidazolium acetate produced competitive results when followed by xylanases before cellulose hydrolysis. These two pretreatment steps helped to improve the accessibility of cellulose to enzymes and enabled cellulosic degradation of up to 99% and a xylose yield of 97.6%
[26][48].
Steam Explosion
This commonly used pretreatment method involves the application of high pressure and temperature, followed by a sudden pressure decrease. This process leads to lignocellulosic structure breakdown
[27][49]. Better biomass disruption was observed when a steam explosion was combined with other pretreatment methods. For example, elephant grass was treated with different concentrations of sulfuric acid to yield around 52% digestible cellulose, while barley straw biomass pretreated with steam explosion and extrusion yielded up to 84% glucan
[28][29][30][31][32][50,51,52,53,54]. However, like dilute acid pretreatment, this pretreatment method results in the production of byproducts that inhibit enzymatic hydrolysis. Thus, a detoxification step is required before enzymatic hydrolysis
[33][55]. The formation of acetic acid, furfural, 5-HMF, and vanillin produced insignificant cellulase inhibition, but formic acid inactivated the enzymes, and this effect increased with the solid loading
[34][56].
Hydrothermal Pretreatment
Hydrothermal pretreatment (HTP) is another example of physicochemical pretreatments. It involves the use of highly pressurized water at high temperatures to disrupt the lignocellulosic structure. HTP can be divided into subcritical and supercritical processes
[27][49]. Due to environmentally friendly conditions, such as no catalyst used, less corrosion, and a high output process, HTP is favored in combination with other pretreatment methods. For example, it has been used in combination with alkaline pretreatment using NaOH, resulting in 86% lignin removal from wheat straw. In addition, it is recognized to be able to enhance the biomass surface area, which enhances the sugar yield in the subsequent steps and enhances the dissolution and recovery of hemicellulose
[35][57].
2.4. Biological Pretreatment
Biological pretreatment involves the use of bacterial and fungal strains, such as
Bacillus sp.,
Trichoderma reesei,
Thermomonospora sp., and
Phanerochaete chrysosporium, to degrade the lignocellulose structure
[36][37][58,59]. Due to the ability of these organisms to release enzymes, such as lignin peroxidase and laccases, lignin is removed from the lignocellulosic structure
[38][60]. The most commonly applied fungus, which is known for its ability to degrade lignocelluloses, is the white-rot fungus. Using
Ceriporiopsis subvermispora to pretreat sugarcane bagasse at 27 °C for 60 days resulted in a 47% sugar yield
[39][61]. Biological pretreatment represents an advance over other pretreatment technology in that it does not require energy input and is environmentally friendly and cost-efficient. However, it is time-consuming and produces low yields; thus, it is infeasible for biorefinery process implementation
[40][62].
3. Hydrolysis
There are several ways in which cellulose can be hydrolyzed to fermentable sugars. Chemical, biological, and other methods such as gamma-ray, electron-beam, and microwave irradiation have been reported. Chemical and biological hydrolysis are the most commonly used due to their feasibility and effectiveness
[41][42][43][63,64,65].
3.1. Chemical Hydrolysis
Chemical hydrolysis involves the use of chemicals such as diluted and concentrated acids. The use of concentrated acid helps to enhance the sugar yield, and such treatments can be performed using lower temperatures than those used in dilute acid hydrolysis. However, acid consumption is high, and further downstream processing, such as detoxification, is required. In addition, this process requires long residence times and recovering acid for reuse is costly. In contrast, dilute acid hydrolysis produces lower yields, requires high temperatures, and generates non-useful byproducts
[44][45][66,67].
3.2. Enzymatic Hydrolysis
The massive application of different cellulases in various fields has attracted attention to their use in bioenergy production. The use of enzymes in those applications helps in reducing the environmental influences and can contribute to enhancing the quality of the production. This is mainly due to the mild conditions required in enzymatic processes, which results in less energy consumption. Moreover, the lower toxins produced, as compared to the chemically catalyzed processes, make enzymatic approaches a good alternative that lowers the required post-treatment steps
[46][14]. However, the high cost of enzymes, associated with their high production cost, remains the main obstacle facing their large-scale application. Therefore, different enzyme-producing companies are leading studies to economically develop enzymes that can be used in various applications, including biofuel production
[46][14].
The main sources of commercial cellulases are
Trichoderma reesei and
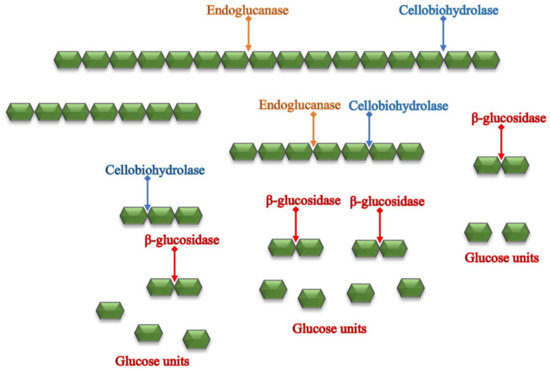
Aspergillus niger [47][68]. Cellulase plays a key role in enzymatic hydrolysis. It is a multi-component system that breaks polymer chains into fermentable sugars, as shown in
Figure 2 [48][69]. It is composed of three types of enzymes: endoglucanase, cellobiohydrolases (exoglucanase), and β-glucosidases. Endoglucanases work synergistically with exoglucanase. Hydrolysis initiates with endoglucanase, which attacks the polymer chain at random sites to create reducing and non-reducing ends. Exoglucanase then acts on those ends to convert them to shorter polysaccharide chains that consist of two glucose units called cellobiose. The last component, β-glucosidase, breaks down cellobiose from the mid-point to produce two glucose units. The latter is considered the rate-limiting step for the hydrolysis reaction because the sensitivity of the enzymes toward the end product, glucose, leads to product inhibition
[49][70]. Studies have shown that each cellulase-producing microorganism lacks one or more types of cellulase, which leads to inefficient hydrolysis.
A. niger and
Trichoderma atroviride are mostly β-glucosidase producers and lack the other two cellulases
[49][70]. Thus, the use of different cellulase recipes from different sources is critical to enhancing the conversion rate. It has been reported that the catalytic activities of commercial cellulases derived from
T. reesei and
A. niger can be improved via the addition of crude cellulases from five different fungal strains, namely
Chaetomium thermophilum, Thielavia terrestris, Thermoascus aurantiacus, Corynascus thermophilus, and
Myceliophthora thermophile, when hydrolyzing pretreated barley straw
[50][71].
Figure 2. A schematic diagram of the bioconversion of lignocellulosic biomass to fermentable glucose [51]. A schematic diagram of the bioconversion of lignocellulosic biomass to fermentable glucose [72].
4. Enzyme Kinetics and Modeling
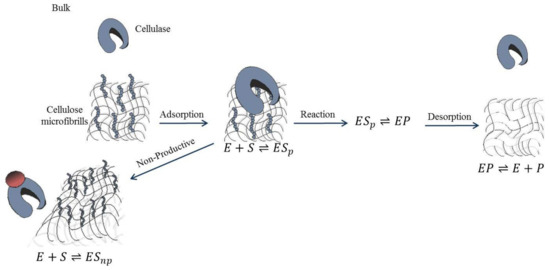
The mechanism of the enzymatic hydrolysis of cellulose is similar to other enzymatic reactions.
Figure 34 illustrates the mechanism and kinetics of each step. In the first step, the enzyme is adsorbed onto the substrate surface. After this, two pathways are possible; the enzyme binds either to an active site, denoted as productive binding, or to a non-active site, denoted as non-productive binding. In the former pathway, an enzyme–substrate complex is formed and can proceed to the catalytic reaction step, during which the glycosidic bond is broken. The enzyme-product complex is formed and then separated to release the product from the enzyme, and the enzyme active site is free again for another binding. Therefore, the rate of the catalytic reaction is directly proportional to the rate of substrate productive binding to the active sites
[52][73]. However, if the substrate is adsorbed via a non-productive pathway, the substrate acts as an inhibitor, and the catalytic process is inhibited. No product is formed in this pathway, and the enzyme is inactive
[52][73]. This demonstrates that the substrate surface changes dynamically as enzymatic hydrolysis progresses
[53][74]. Cellulose is composed of hydrolyzable and non-hydrolyzable (inert) parts. At the enzyme–substrate surface, the enzyme breaks down cellulose leaving the inert at that layer, proceeding to the next layer, which also contains cellulose and inert
[53][74]. As the reaction continues, the enzyme is adsorbed deeper into the substrate, shrinking the available substrate surface area and thus limiting the substrate available for reaction.
Figure 34.
Reaction mechanism of enzymatic hydrolysis of cellulose.
Various cellulase kinetic mechanisms have been proposed by researchers who have sought to understand the process fully. The mechanism shown in Equation (1) was proposed to explain the burst phase for a soluble substrate and non-processive enzyme, which cleaves cellulose randomly and non-processively. When the processive action of an enzyme occurs, the enzyme binds to the cellulose and cleaves it continuously for multiple cycles before it dissociates
[54][75].
The model suggests that an enzymatic reaction produces two products. The first,
P1, is produced rapidly at the beginning of the reaction when
k1 and
k2 are larger than
k3. The enzyme then accumulates as an
EP2 complex, which dissociates slowly over time to produce
P2. This can be proven by monitoring the concentrations of
P1 and
P2 as functions of time.
P1 increases during the first stage of the reaction, before the reaction reaches steady-state
[54][55][56][75,76,77]. However, key factors that affect the enzyme mechanisms, such as product inhibition, are not considered.
To account for the processive action of the enzyme, Equation (1) was modified to include multiple cycles of the catalytic action of cellobiohydrolase, as shown in Equation (2).
In this model of cellulose cleavage, the enzyme cleaves the cellulose strand in consecutive steps in a processive manner and goes through many cycles of consecutive reactions before it dissociates. Hydrolysis initiates via the binding of cellobiohydrolase,
E, to a cellulose strand that contains n cellobiose units,
Cn, to form an enzyme–substrate complex,
ECn. After this, it can either produce one cellobiose unit,
C, with a rate constant k
2 or dissociate back to the enzyme and cellulose with the dissociation rate constant
k3. The reaction continues via the same steps but with shorter cellulose strands in each cycle and with the possibility of enzyme dissociation. After multiple cycles of cellobiose unit cleavage, the desorption rate slows because inhibition by cellobiose leads to enzyme–substrate complex (
ECx) accumulation. This occurs because
k2 is larger than
k3, and a further decline in kinetics is observed
[54][55][56][75,76,77].
The drawback of this model is that it assumes a constant substrate concentration. This is valid only at the beginning of the reaction when the substrate concentration is higher than the enzyme concentration. Hence, the accessibility and affinity of the enzyme to bind to the remainder of the cellulose strand is not considered
[57][78]. In addition, the inhibition effect is considered only with respect to cellobiose; mono-sugars are not considered. This model has been modified further to consider enzyme deactivation via irreversible binding to the substrate, as in Equation (3).