The atrial natriuretic peptide (ANP), a cardiovascular hormone, plays a pivotal role in the homeostatic control of blood pressure, electrolytes, and water balance and is approved to treat congestive heart failure. In addition, there is a growing realization that ANPs might be related to immune response and tumor growth. The anti-inflammatory and immune-modulatory effects of ANPs in the tissue microenvironment are mediated through autocrine or paracrine mechanisms, which further suppress tumorigenesis. In cancers, ANPs show anti-proliferative effects through several molecular pathways. Furthermore, ANPs attenuate the side effects of cancer therapy. Therefore, ANPs act on several hallmarks of cancer, such as inflammation, angiogenesis, sustained tumor growth, and metastasis.
- atrial natriuretic peptide
- inflammation
- anti-cancer agent
1. Modulation of Inflammation and Anti-Tumor Effects
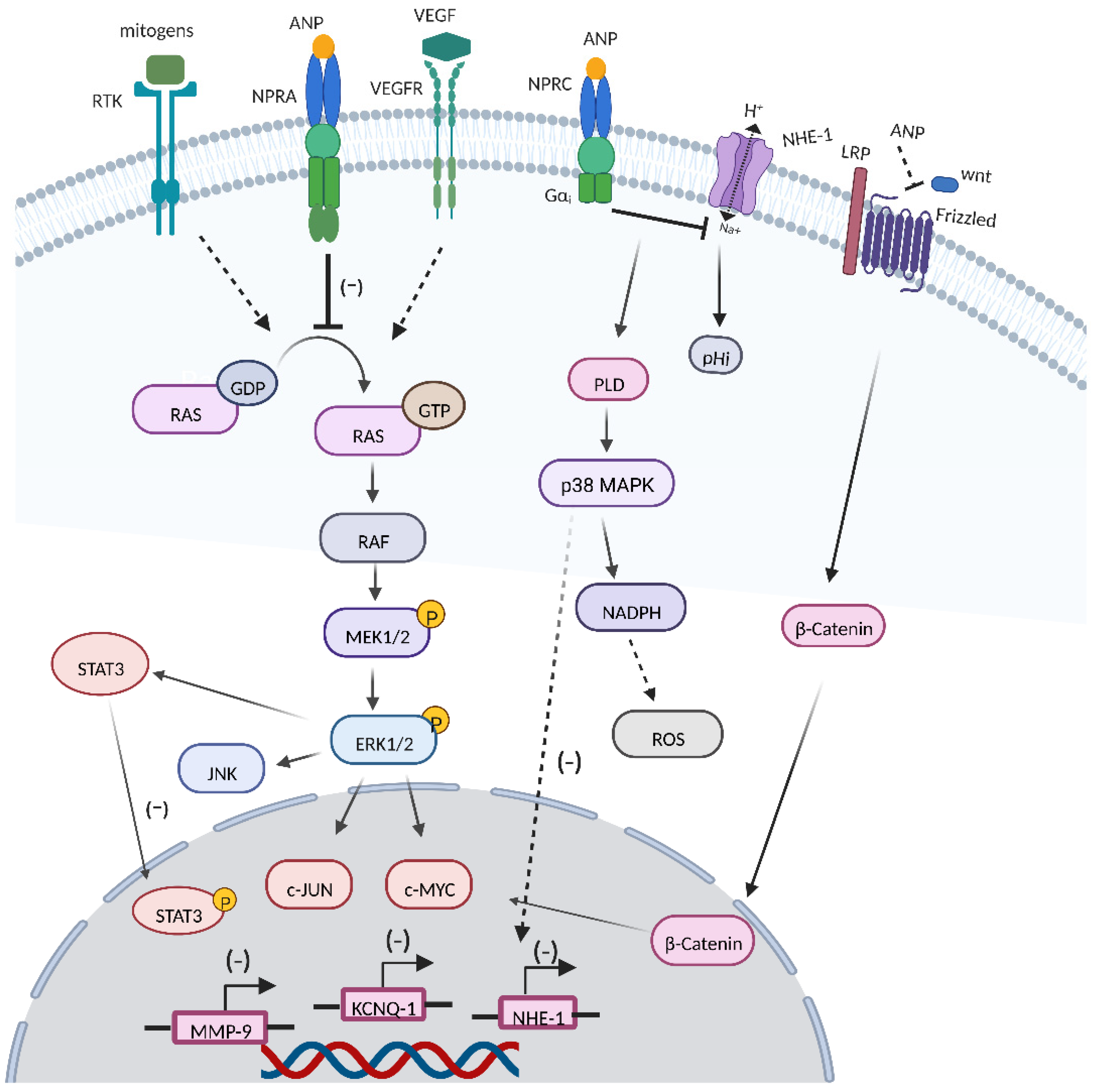
2. RAS-MEK1/2-ERK1/2 Kinase Cascade
3. ANPs Interact with Other Transcription Factors and Cell Signaling Systems
3.1. VEGF
3.2. Wnt/β-Catenin Signaling Cascade
3.3. JNK and JAK/STATs Signaling
3.4. ROS Production
3.5. KCNQ1 Expression
3.6. MMP9 Expression
References
- Shalapour, S.; Karin, M. Immunity, inflammation, and cancer: An eternal fight between good and evil. J. Clin. Investig. 2015, 125, 3347–3355.
- Balkwill, F.; Mantovani, A. Inflammation and cancer: Back to Virchow? Lancet 2001, 357, 539–545.
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674.
- Grivennikov, S.I.; Greten, F.R.; Karin, M. Immunity, inflammation, and cancer. Cell 2010, 140, 883–899.
- Khansari, N.; Shakiba, Y.; Mahmoudi, M. Chronic inflammation and oxidative stress as a major cause of age-related diseases and cancer. Recent Pat. Inflamm. Allergy Drug Discov. 2009, 3, 73–80.
- Vollmar, A.M. The role of atrial natriuretic peptide in the immune system. Peptides 2005, 26, 1086–1094.
- Vellaichamy, E.; Khurana, M.L.; Fink, J.; Pandey, K.N. Involvement of the NF-kappa B/matrix metalloproteinase pathway in cardiac fibrosis of mice lacking guanylyl cyclase/natriuretic peptide receptor A. J. Biol. Chem. 2005, 280, 19230–19242.
- Vellaichamy, E.; Kaur, K.; Pandey, K.N. Enhanced activation of pro-inflammatory cytokines in mice lacking natriuretic peptide receptor-A. Peptides 2007, 28, 893–899.
- Subramanian, V.; Vellaichamy, E. Atrial natriuretic peptide (ANP) inhibits DMBA/croton oil induced skin tumor growth by modulating NF-κB, MMPs, and infiltrating mast cells in swiss albino mice. Eur. J. Pharmacol. 2014, 740, 388–397.
- Archer, M.; Dogra, N.; Kyprianou, N. Inflammation as a Driver of Prostate Cancer Metastasis and Therapeutic Resistance. Cancers 2020, 12, 2984.
- Mezzasoma, L.; Costanzi, E.; Scarpelli, P.; Talesa, V.N.; Bellezza, I. Extracellular Vesicles from Human Advanced-Stage Prostate Cancer Cells Modify the Inflammatory Response of Microenvironment-Residing Cells. Cancers 2019, 11, 1276.
- Mezzasoma, L.; Talesa, V.N.; Costanzi, E.; Bellezza, I. Natriuretic Peptides Regulate Prostate Cells Inflammatory Behavior: Potential Novel Anticancer Agents for Prostate Cancer. Biomolecules 2021, 11, 794.
- Funaki, S.; Sawabata, N.; Nakagiri, T.; Shintani, Y.; Inoue, M.; Kadota, Y.; Minami, M.; Okumura, M. Novel approach for detection of isolated tumor cells in pulmonary vein using negative selection method: Morphological classification and clinical implications. Eur. J. Cardiothorac. Surg. 2011, 40, 322–327.
- Giavazzi, R.; Foppolo, M.; Dossi, R.; Remuzzi, A. Rolling and adhesion of human tumor cells on vascular endothelium under physiological flow conditions. J. Clin. Investig. 1993, 92, 3038–3044.
- Koch, M.; Kienle, P.; Hinz, U.; Antolovic, D.; Schmidt, J.; Herfarth, C.; von Knebel Doeberitz, M.; Weitz, J. Detection of hematogenous tumor cell dissemination predicts tumor relapse in patients undergoing surgical resection of colorectal liver metastases. Ann. Surg. 2005, 241, 199–205.
- McDonald, B.; Spicer, J.; Giannais, B.; Fallavollita, L.; Brodt, P.; Ferri, L.E. Systemic inflammation increases cancer cell adhesion to hepatic sinusoids by neutrophil mediated mechanisms. Int. J. Cancer 2009, 125, 1298–1305.
- Nojiri, T.; Inoue, M.; Yamamoto, K.; Maeda, H.; Takeuchi, Y.; Funakoshi, Y.; Okumura, M. Effects of low-dose human atrial natriuretic peptide for preventing post-operative cardiopulmonary complications in elderly patients undergoing pulmonary resection for lung cancer. Eur. J. Cardiothorac. Surg. 2012, 41, 1330–1334.
- Nojiri, T.; Yamamoto, K.; Maeda, H.; Takeuchi, Y.; Funakoshi, Y.; Inoue, M.; Okumura, M. Effect of low-dose human atrial natriuretic peptide on postoperative atrial fibrillation in patients undergoing pulmonary resection for lung cancer: A double-blind, placebo-controlled study. J. Thorac. Cardiovasc. Surg. 2012, 143, 488–494.
- Nojiri, T.; Arai, M.; Suzuki, Y.; Kumazoe, M.; Tokudome, T.; Miura, K.; Hino, J.; Hosoda, H.; Miyazato, M.; Okumura, M.; et al. Transcriptome analysis reveals a role for the endothelial ANP-GC-A signaling in interfering with pre-metastatic niche formation by solid cancers. Oncotarget 2017, 8, 65534–65547.
- Kong, X.; Wang, X.; Xu, W.; Behera, S.; Hellermann, G.; Kumar, A.; Lockey, R.F.; Mohapatra, S.; Mohapatra, S.S. Natriuretic peptide receptor a as a novel anticancer target. Cancer Res. 2008, 68, 249–256.
- Mallela, J.; Ravi, S.; Jean Louis, F.; Mulaney, B.; Cheung, M.; Sree Garapati, U.; Chinnasamy, V.; Wang, C.; Nagaraj, S.; Mohapatra, S.S.; et al. Natriuretic peptide receptor A signaling regulates stem cell recruitment and angiogenesis: A model to study linkage between inflammation and tumorigenesis. Stem Cells 2013, 31, 1321–1329.
- Davies, H.; Bignell, G.R.; Cox, C.; Stephens, P.; Edkins, S.; Clegg, S.; Teague, J.; Woffendin, H.; Garnett, M.J.; Bottomley, W.; et al. Mutations of the BRAF gene in human cancer. Nature 2002, 417, 949–954.
- Sebolt-Leopold, J.S. Advances in the development of cancer therapeutics directed against the RAS-mitogen-activated protein kinase pathway. Clin. Cancer Res. 2008, 14, 3651–3656.
- Sun, Y.; Eichelbaum, E.J.; Wang, H.; Vesely, D.L. Atrial natriuretic peptide and long acting natriuretic peptide inhibit MEK 1/2 activation in human prostate cancer cells. Anticancer Res. 2007, 27, 3813–3818.
- Medema, R.H.; de Vries-Smits, A.M.; van der Zon, G.C.; Maassen, J.A.; Bos, J.L. Ras activation by insulin and epidermal growth factor through enhanced exchange of guanine nucleotides on p21ras. Mol. Cell. Biol. 1993, 13, 155–162.
- Vesely, D.L. Natriuretic peptides’ metabolic targets for treatment of cancer. J. Investig. Med. 2013, 61, 816–822.
- Apte, R.S.; Chen, D.S.; Ferrara, N. VEGF in Signaling and Disease: Beyond Discovery and Development. Cell 2019, 176, 1248–1264.
- Kerbel, R.S. Tumor angiogenesis. N. Engl. J. Med. 2008, 358, 2039–2049.
- Ribatti, D.; Solimando, A.G.; Pezzella, F. The Anti-VEGF(R) Drug Discovery Legacy: Improving Attrition Rates by Breaking the Vicious Cycle of Angiogenesis in Cancer. Cancers 2021, 13, 3433.
- Pedram, A.; Razandi, M.; Levin, E.R. Natriuretic peptides suppress vascular endothelial cell growth factor signaling to angiogenesis. Endocrinology 2001, 142, 1578–1586.
- Levin, E.R.; Gardner, D.G.; Samson, W.K. Natriuretic peptides. N. Engl. J. Med. 1998, 339, 321–328.
- Kook, H.; Itoh, H.; Choi, B.S.; Sawada, N.; Doi, K.; Hwang, T.J.; Kim, K.K.; Arai, H.; Baik, Y.H.; Nakao, K. Physiological concentration of atrial natriuretic peptide induces endothelial regeneration in vitro. Am. J. Physiol. Heart Circ. Physiol. 2003, 284, H1388–H1397.
- Nguyen, J.P.; Frost, C.D.; Lane, M.L.; Skelton, W.I.; Skelton, M.; Vesely, D.L. Novel dual inhibitors of vascular endothelial growth factor and VEGFR2 receptor. Eur. J. Clin. Investig. 2012, 42, 1061–1067.
- Mao, G.; Zheng, S.; Li, J.; Liu, X.; Zhou, Q.; Cao, J.; Zhang, Q.; Zheng, L.; Wang, L.; Qi, C. Glipizide Combined with ANP Suppresses Breast Cancer Growth and Metastasis by Inhibiting Angiogenesis through VEGF/VEGFR2 Signaling. Anticancer Agents Med. Chem. 2022, 22, 1735–1741.
- Nakao, Y.; Yamada, S.; Yanamoto, S.; Tomioka, T.; Naruse, T.; Ikeda, T.; Kurita, H.; Umeda, M. Natriuretic peptide receptor A is related to the expression of vascular endothelial growth factors A and C, and is associated with the invasion potential of tongue squamous cell carcinoma. Int. J. Oral Maxillofac. Surg. 2017, 46, 1237–1242.
- Barzegar, B.A.; Talaie, Z.; Jusheghani, F.; Łos, M.J.; Klonisch, T.; Ghavami, S. Wnt and PI3K/Akt/mTOR Survival Pathways as Therapeutic Targets in Glioblastoma. Int. J. Mol. Sci. 2022, 23, 1353.
- Vesely, D.L. Cardiac hormones for the treatment of cancer. Endocr. Relat. Cancer 2013, 20, R113–R125.
- Kanda, F.; Sarnacki, P.; Arieff, A.I. Atrial natriuretic peptide inhibits amiloride-sensitive sodium uptake in rat brain. Am. J. Physiol. 1992, 263 Pt 2, R279–R283.
- Ricci, R.; Baldini, P.; Bogetto, L.; De Vito, P.; Luly, P.; Zannetti, A.; Incerpi, S. Dual modulation of Na/H antiport by atrial natriuretic factor in rat aortic smooth muscle cells. Am. J. Physiol. 1997, 273 Pt 1, C643–C652.
- Serafino, A.; Moroni, N.; Psaila, R.; Zonfrillo, M.; Andreola, F.; Wannenes, F.; Mercuri, L.; Rasi, G.; Pierimarchi, P. Anti-proliferative effect of atrial natriuretic peptide on colorectal cancer cells: Evidence for an Akt-mediated cross-talk between NHE-1 activity and Wnt/β-catenin signaling. Biochim. Biophys. Acta 2012, 1822, 1004–1018.
- Kumar, A.; Singh, U.K.; Kini, S.G.; Garg, V.; Agrawal, S.; Tomar, P.K.; Pathak, P.; Chaudhary, A.; Gupta, P.; Malik, A. JNK pathway signaling: A novel and smarter therapeutic targets for various biological diseases. Future Med. Chem. 2015, 7, 2065–2086.
- Vesely, D.L. Heart Peptide Hormones: Adjunct and Primary Treatments of Cancer. Anticancer Res. 2016, 36, 5693–5700.
- Yu, H.; Lee, H.; Herrmann, A.; Buettner, R.; Jove, R. Revisiting STAT3 signalling in cancer: New and unexpected biological functions. Nat. Rev. Cancer 2014, 14, 736–746.
- Galadari, S.; Rahman, A.; Pallichankandy, S.; Thayyullathil, F. Reactive oxygen species and cancer paradox: To promote or to suppress? Free Radic. Biol. Med. 2017, 104, 144–164.
- Tang, F.; Liu, S.; Li, Q.Y.; Yuan, J.; Li, L.; Wang, Y.; Yuan, B.F.; Feng, Y.Q. Location analysis of 8-oxo-7,8-dihydroguanine in DNA by polymerase-mediated differential coding. Chem. Sci. 2019, 10, 4272–4281.
- Tang, F.; Yuan, J.; Yuan, B.F.; Wang, Y. DNA-Protein Cross-Linking Sequencing for Genome-Wide Mapping of Thymidine Glycol. J. Am. Chem. Soc. 2022, 144, 454–462.
- Trachootham, D.; Alexandre, J.; Huang, P. Targeting cancer cells by ROS-mediated mechanisms: A radical therapeutic approach? Nat. Rev. Drug Discov. 2009, 8, 579–591.
- NavaneethaKrishnan, S.; Rosales, J.L.; Lee, K.Y. Targeting Cdk5 for killing of breast cancer cells via perturbation of redox homeostasis. Oncoscience 2018, 5, 152–154.
- Rashed, H.M.; Sun, H.; Patel, T.B. Atrial natriuretic peptide inhibits growth of hepatoblastoma (HEP G2) cells by means of activation of clearance receptors. Hepatology 1993, 17, 677–684.
- Baldini, P.M.; De Vito, P.; Vismara, D.; Bagni, C.; Zalfa, F.; Minieri, M.; Di Nardo, P. Atrial natriuretic peptide effects on intracellular pH changes and ROS production in HEPG2 cells: Role of p38 MAPK and phospholipase D. Cell. Physiol. Biochem. 2005, 15, 77–88.
- Baldini, P.M.; De Vito, P.; Antenucci, D.; Vismara, D.; D’Aquilio, F.; Luly, P.; Zalfa, F.; Bagni, C.; Di Nardo, P. Atrial natriuretic peptide induces cell death in human hepatoblastoma (HepG2) through the involvement of NADPH oxidase. Cell Death Differ. 2004, 11 (Suppl. 2), S210–S212.
- Kiemer, A.K.; Gerbes, A.L.; Bilzer, M.; Vollmar, A.M. The atrial natriuretic peptide and cGMP: Novel activators of the heat shock response in rat livers. Hepatology 2002, 35, 88–94.
- De Vito, P. Atrial natriuretic peptide: An old hormone or a new cytokine? Peptides 2014, 58, 108–116.
- Zhanping, W.; Xiaoyu, P.; Na, C.; Shenglan, W.; Bo, W. Voltage-gated K+ channels are associated with cell proliferation and cell cycle of ovarian cancer cell. Gynecol. Oncol. 2007, 104, 455–460.
- Spitzner, M.; Ousingsawat, J.; Scheidt, K.; Kunzelmann, K.; Schreiber, R. Voltage-gated K+ channels support proliferation of colonic carcinoma cells. FASEB J. 2007, 21, 35–44.
- Zhang, J.; Zhao, Z.; Zu, C.; Hu, H.; Shen, H.; Zhang, M.; Wang, J. Atrial natriuretic peptide modulates the proliferation of human gastric cancer cells via KCNQ1 expression. Oncol. Lett. 2013, 6, 407–414.
- Zhang, J.; Qu, J.; Yang, Y.; Li, M.; Zhang, M.; Cui, X.; Zhang, J.; Wang, J. Impact of NPR-A expression in gastric cancer cells. Int. J. Clin. Exp. Med. 2014, 7, 3209–3214.
- Mondal, S.; Adhikari, N.; Banerjee, S.; Amin, S.A.; Jha, T. Matrix metalloproteinase-9 (MMP-9) and its inhibitors in cancer: A minireview. Eur. J. Med. Chem. 2020, 194, 112260.
- Li, C.H.; Liu, M.; Pan, L.H.; Sun, Y. ANP reduced Hedgehog signaling-mediated activation of matrix metalloproteinase-9 in gastric cancer cell line MGC-803. Gene 2020, 762, 145044.