23. Gut Microbiota in Infants and Toddlers under Standard Diets
2.1. Breast Milk vs. Formula Milk and Gut Microbiota
3.1. Breast Milk vs. Formula Milk and Gut Microbiota
Breast milk, composed of lactose, lipids, proteins, and more than 200 different human milk oligosaccharides (HMOs)
[17][18], is recommended as the first nutrition during infancy, providing all the necessary nutrients to support growth and development
[18][19][20][19,20,21]. HMOs are not hydrolyzed by gastrointestinal saccharolytic enzymes; thus, they reach the colon intact, where they serve as metabolic substrates for gut bifidobacteria
[8]. Some bifidobacterial species (e.g.,
B. bifidum,
B. breve and subsp.
infantis) possess specific membrane transporters and saccharolytic enzymes that are capable of cleaving and internalizing HMOs, producing lactate, acetate, formate, and 1,2-propanediol
[21][22]. The end products of HMOs’ metabolism have potential beneficial host-health effects. Indeed, acetate and lactate have been reported to suppress the growth of opportunistic pathogens belonging to
Clostridiaceae,
Enterobacteriaceae, and
Staphylococcaceae [8][22][8,23]. The lack of
Bifidobacterium spp. has been instead associated with immune dysregulation
[23][24], asthma
[24][25], and autoimmune diseases
[25][26]. Interestingly, Alcon-Giner et al. conducted a longitudinal study to evaluate the gut microbiota composition in two groups of preterm infants (101 orally supplemented with
Bifidobacterium and
Lactobacillus -Bif/Lacto- species and 133 non-supplemented). They observed enrichment in
Bifidobacterium and a lower abundance of opportunistic pathogens in the Bif/Lacto group
[26][27]. Moreover, fecal acetate positively correlated with
Bifidobacterium relative abundance
[26][27].
Overall, HMO and lactose promote the GI microbiota development and stimulated the infant immune system through growth factors, cytokines and immunoglobulins, with a positive impact on growth, neurodevelopment and metabolism
[27][28].
Exclusively formula-fed (FF) infants possess different microbiota, characterized by a lower abundance of HMO-utilizing
Bifidobacterium species and increased
Clostridium species (e.g.,
C. difficile and
C. perfringens) and
Enterobacteriaceae (e.g.,
Escherichia coli)
[28][29][29,30]. These observations seem to correlate with the lack of HMOs and the higher protein content in milk formulas
[8]. Recently, Berger et al., in a double-blinded randomized controlled trial, evaluated whether supplementation of infant formula with HMOs, (specifically 2′-fucosyl-lactose-2′FL- and lacto-N-neotetraose-LNnT) could influence the gut microbiota
[30][31]. Notably, recent studies have evaluated the effect of infant formula supplemented with synthetic HMOs (e.g., 2′FL and LNnT) in order to mimic a BF-like gut metabolism and microbial colonization
[31][32]. Moreover, the researchers compared HMO-supplemented and un-supplemented formula groups with BF infants
[30][32][31,33]. The intervention group led to a microbiota pattern that was more similar to that of the breast-fed (BF) infants, with higher levels of
Bifidobacterium spp. and a lower relative abundance of
Clostridiaceae and
Enterobacteriaceae, compared with infants fed with un-supplemented formula
[30][31].
Many infant formulas are, nowadays, supplemented with fructo-oligo-saccharides (FOS) and/or galacto-oligosaccharides (GOS), although these are not selected as maternal HMOs as they can be utilized by most
Bifidobacterium species (e.g.,
B. adolescentis and
B. catenulatum, typical of adulthood)
[28][29][29,30]. Studies comparing the fecal metabolome in exclusively BF and exclusively FF infants (even when the formula contains galacto-oligosaccharides) show that proteolytic rather than saccharolytic metabolism dominates in the FF gut
[33][34][34,35]. Indeed, in BF infants, remnants of HMO metabolism are mainly present, while in FF infants, the main observed products are derived from protein fermentation
[33][34][34,35]. Thus, the authors evaluated the intestinal fermentation capability, which might be influenced by early diet
[33][34]. The different fecal microbiota were also related to the differences in the fecal water % and in the concentration of microbial by-products in the stool
[33][34]. In FF infants, limited oligosaccharides and high protein levels were present
[33][34]. The difference in terms of fecal microbial taxonomic profiles of the milk fat globule membrane (MFGM) supplemented with experimental formula (EF) and standard formula (SF) was moderate, with respect to the difference between BF and FF infants
[33][34]. However, to confirm these results, further studies are needed.
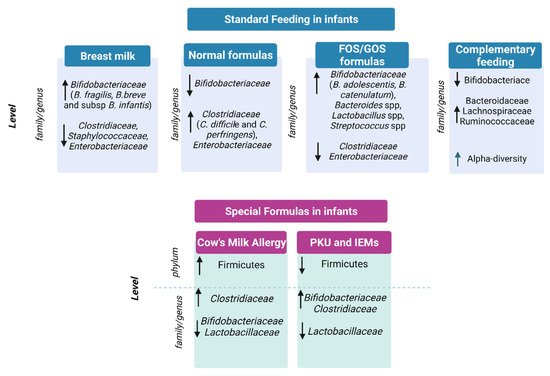
In conclusion, breastfeeding, mainly due to the presence of HMOs, promotes the colonization of beneficial bifidobacteria and is able to support immune system development and to prevent infections, as depicted in
Figure 1 [8][22][35][36][37][8,23,36,37,38]. On the contrary, milk formulas have been correlated to the growth of opportunistic pathogens and to proteolytic gut metabolism, potentially causing adverse health effects
[38][39][40][41][39,40,41,42].
Figure 1. Gut microbiota in standard feeding vs. special formula-fed infants. ↓= decrease in family/genus or phylum; ↑ = increase in genus/family or phylum.
2.2. Gut Microbiota and Complementary Feeding
3.2. Gut Microbiota and Complementary Feeding
Complementary feeding is a period characterized by a shift from an exclusive milk diet to a mixed diet, including food consumption and it goes along with a phase of remarkable changes in the gut microbiota, of which the most relevant is the rapid decline in HMO-degrading
Bifidobacterium species
[42][43]. Moreover, alpha diversity increases and higher levels of
Bacteroidaceae,
Lachnospiraceae, and
Ruminococcaceae species are present. This mirrors the higher diversity and complexity of the diet, due to the introduction of fibers (from fruits, vegetables and cereals) and new protein sources (as meats, dairy products, and legumes)
[8][13][8,14].
Coherently, Differding et al. associated
Bilophila wadsworthia and
Roseburia spp. with early complementary feeding practice (e.g., consumption of solids or non-water/formula liquids at or before 3 months)
[43][44]. Leong et al., in a randomized controlled trial, compared the gut microbiota composition of children weaned with a “baby-led” approach of complementary feeding with traditional spoon-fed infants
[44][45]. The baby-led weaning approach was associated with lower alpha diversity and lower levels of
Lachnospiraceae (
Roseburia facies and
Eubacterium rectale) and
Ruminococcaceae (
Faecalibacterium prausnitzii)
[44][45]. The multinational European INFABIO study (
n = 605 infants) found that early introduction of complementary foods was associated with a decrease in bifidobacteria (sp. unknown)
[45][46]. Specifically, the study confirmed that the initial feeding method influenced the relative abundance of the
Clostridium leptum group,
Clostridioides difficile and
C perfringens, and
bifidobacteria dominated the feces of initially breast-fed infants
[45][46].
Some complementary foods, such as rye bread, cheese, and meat products, have been specifically correlated to this increase in diversification of the infant gut microbiota
[46][47]. Importantly, complementary feeding progression has also been correlated to higher levels of several
Lachnospiraceae and
Ruminococcaceae, and to a lower abundance in
Bifidobacterium spp.
[13][14]. This underlines the transition from a bifidobacteria-rich gut community toward a more diverse gut microbial community, rich in butyrate-, propionate-, and BCFA-producing bacteria
[13][14].
Concerning the different food introduced, meat causes significantly increased levels of genera that belong to
Lachnospiraceae, which are potential short-chain fatty acid producers with respect to dairy
[47][48]. Recently, a comparison between refined grain cereal products and whole grain cereal products as the first complementary food showed a significant increase in
Bacteroides and
Lachnoclostridium (Lachnospiraceae family) and a decrease in
Escherichia (
Enterobacteriaceae family) in the whole grain cereal group
[46][47].
34. Gut Microbiota in Infants under Special Diets
3.1. Special Formulas for the Treatment of Cow’s Milk Allergy
4.1. Special Formulas for the Treatment of Cow’s Milk Allergy
Cow’s milk protein allergy (CMA) is the most common food allergy in the pediatric population
[48][49]. The use of extensively hydrolyzed formulas (eHF) is recommended as the first choice for the treatment of infants and children with confirmed CMA
[49][50]. Amino acid-based formulas (AAFs) are also available and are considered the safest dietary strategy for infants with severe CMA
[50][51][51,52]. Hydrolyzed formulas minimize antigen contact compared to whole-protein formulas, but due to the presence of sparse small peptide fragments, it also may favor immune tolerance by different mechanisms, acting on barrier reinforcement, neuro-immune pathways, and on the resident microbial community
[52][53]. Indeed, the peptides derived from protein hydrolysates can act locally in the gut, modulating the microbiota. Dysbiosis induces functional alterations that result in aberrant Th2 responses toward an allergic, rather than a tolerogenic, response
[53][54]. Conversely, recent data suggest that the use of AAF, the only totally non-allergenic formulas, exclusively providing nitrogen equivalent proteins as free amino acids, may delay the acquisition of immune tolerance
[54][55], since they are completely devoid of allergenic antigenic epitopes
[49][55][50,56]. Paparo et al. showed the absence of AAF-related effects on the intestinal barrier, Th1/Th2 cytokine response, and regulatory T cells (Treg) activation in vitro
[56][57]. The onset of oral tolerance occurs during the critical early stages of immune development and it depends not only on genetic factors, but also on environmental factors, including appropriate intestinal bacterial colonization
[57][58]. Research on the development of oral tolerance has recently focused on the influence of host–microbe interactions on immune function, in particular on the ability of specific gut microbes to promote tolerance to food antigens
[58][59][60][59,60,61]. In this respect, it has been reported that supplementation with
Lactobacilllus rhamnosus GG (in an extensively hydrolyzed casein formula, eHCF) in infants with CMA stimulates immune tolerance
[54][61][62][55,62,63], and some eHFs were supplemented with specific probiotic strains, which in turn may act on immunomodulation, in addition to the small peptides. Berni Canani et al. found that dietary management with an eHCF formula supplemented with the probiotic
Lactobacillus rhamnosus GG (LGG) results in a higher rate of tolerance acquisition in children with IgE-mediated CMA, compared with non-supplemented eHCF or non-casein-based formulas
[54][55]. In addition, in a well-characterized population of children with IgE-mediated CMA, it was shown that eHCF + LGG is superior to eHCF alone in preventing other allergic manifestations over a 36-month period
[63][64]. The gut microbiota produces metabolites, such as SCFAs, beneficial for the colonic environment, which participate in improving barrier function, increasing Treg cells, and protecting against pathogenic bacteria colonization
[64][65][65,66]. Another of the hypothesized mechanisms for an increased immune tolerance is the enrichment of butyrate-producing bacteria, as butyrate stimulates the development of Treg. Treg cells play a pivotal role in the development of oral tolerance
[66][67][68][67,68,69] and their reduction has been associated with an allergic phenotype
[69][70][70,71]. Ruohtula and colleagues found that an increase in highly activated Treg cells is associated with colonization of butyrate-producing bacilli and suggested a narrow window of opportunity (from birth to 3 months of age) for primary prevention of atopic diseases
[71][72]. eHFs favour butyrate production, as highlighted by Car RennKok et al., who compared healthy term breastfed infants vs. infants fed with eHF or AAF. The authors found significant differences between the groups in beta-diversity at day 60, and a similar gut microbiota composition in the formula groups compared to the breastfed group. The relative abundance of
Bifidobacterium spp. increased over time and was significantly enriched at day 60 in the human milk group. In contrast, a significant increase in Firmicutes members was found at day 60 in the formula groups, as well as butyrate-producing species in the eHF group. Butyrate increased significantly from baseline to day 60 in the eHF group and it was significantly higher in the study formula groups than in the human milk at day 60
[72][73]. Berni Canani et al. performed gut microbiota and oligotyping analysis on stool samples collected from healthy infants and infants with CMA before and after treatment with eHCF with or without LGG supplementation. They found that the infants who became tolerant early showed a significant increase in fecal butyrate levels and that eHCF + LGG promotes tolerance in infants with CMA, in part, by influencing the structure of the gut bacterial community at the strain level
[62][63]. The same authors showed that children with non-IgE-mediated CMA had significantly lower fecal butyrate concentrations than healthy controls at the baseline, and dietary regimes (eHCF or eHCF + LGG) were associated with a significant increase in butyrate concentrations, especially in children treated with eHCF + LGG. In particular, the treatment with eHCF + LGG appears to restore the composition and structure of the subgenus
Bacteroides, which shows similar diversity to that of healthy controls. Treatment with eHCF + LGG significantly increased butyrate production, and this change correlated with the enrichment of potential SCFA producers and selected
Bacteroides oligotypes
[73][74].
Besides probiotics, hydrolyzed formulas were also supplemented with oligosaccharides
[74][75]. The effects of a formula supplemented with 2 HMOs-2′fucosyllactose (2′FL) and 0.5 g/L lacto-N-neotetraose (LNnT) on infant growth, tolerance and morbidity have been evaluated vs. non-supplemented formula milk
[74][75]. The authors observed a similar weight gain in the two groups at 4 months, and similar digestive symptoms and behavioral patterns between the groups
[75][76]. The safety and hypoallergenicity of an eHF supplemented with 2′FL and LNnT compared with a standard eHF were successfully assessed by Nowak-Wegrzyn and coworkers
[76][77].
The possible influence of eHF formulas in promoting butyrate-producing species, and consequently butyrate production itself, is of great interest in the development of oral tolerance in infants with CMA. In addition, the role of probiotic-supplemented formulas, particularly LGGs, in the acquisition of immune tolerance is emerging. Future studies are required to elucidate the role of both synbiotics and HMOs in the pediatric population with CMA.
3.2. Special Formula for Functional Gastrointestinal Symptoms
4.2. Special Formula for Functional Gastrointestinal Symptoms
Gastrointestinal symptoms, including frequent regurgitation, vomiting, and colicky, are very common in infants, and represent the most frequent cause of formula change in the first six months of age.
These symptoms can overlap non-IgE CMA ones, making it difficult to distinguish between them in clinical practice. However, it is very important to differentiate between these conditions as CMA is confirmed by oral food challenge only in a small percentage of infants
[77][81] and it requires the use of an eHF, according to the guidelines
[49][50]. A clinical score known as CoMiSS (cow’s milk related symptoms score) has been developed as a clinical tool aimed at increasing the awareness of health care professionals in identifying clinical manifestations possibly related to cow’s milk (CM) intake
[78][82]. In a study comparing the effects of two eHF supplemented with probiotics in infants with suspected CMA, an improvement in all the items was observed after dietary intervention
[79][83]. The observed effects on intestinal microbes were, as expected, an increase in bifidobacteria in the group with eHF supplemented with bifidobacteria, and enrichment in lactobacilli in the group with eHF supplemented with lactobacilli. Both formulas were equally effective and safe.
The effective response to hydrolyzed formulas in both functional gastrointestinal disorders and non-IgE CMA may be due to immunological mechanisms, as well as to improved gastric emptying
[53][80][54,84]. Furthermore, a CM-free diet reduces mast cell infiltration and normalizes immune–nerve interactions and motor function
[53][54].
Comparing a thickened eHF vs. a standard eHF on 77 infants with suspected CMA and bothersome regurgitation or vomiting (more than five episodes per day), a reduction in regurgitation was observed, both upon eHF use (after one month of intervention) and, to a greater extent, upon thickened formula intake
[81][85].
In a French group of 30 infants with proven CMA by a double-blind dietary test (70% with positive IgE tests), the administration of a casein-based thickened eHF significantly decreased both regurgitation and clinical crying scores after only 14 days
[82][86].
3.3. Phenylketonuria and Inborn Errors of Metabolism Formulas
4.3. Phenylketonuria and Inborn Errors of Metabolism Formulas
Studies on the microbiome and its influence in patients with inborn errors of metabolism formulas (IEMs) mostly focused on phenylketonuria (PKU). To date, only one open-label, single-arm intervention pilot study in 2011 has evaluated the tolerance and efficacy of adding a prebiotic oligosaccharide mixture (scGOS/lcFOS) to a protein replacement suitable for children with PKU in 9 infants with PKU diagnosis. At the end of the observation period (8-weeks), all children had microbiota dominated by bifidobacteria, unchanged from the baseline. No child showed abnormally high levels of
Clostridium histolyticum/
lituseburense or potentially pathogenic
Enterobacteriaceae at any time during the study. A significant reduction in median stool pH from the baseline was observed at week 4 (pH from 6.79 to 5.83), but this observation was not confirmed at week 8
[83][99].