1. Introduction
Several examples document the inevitable and continuous interaction between humans and plants, primarily characterized by the human interest in using them for consumption, materials production, medical purposes, decoration, and bioremediation. An example of the most striking human manipulations on the plants’ life and environment is the establishment of urban green spaces, for instance, public parks and/or private residential gardens, with surfaces more or less limited, where plants are grown with objectives often different from those of agriculture
[1]. In fact, if agriculture has aims that are almost exclusively at production, the creation of green areas within urban centers was born as a response to profoundly different problems, such as the demographic increase and the progressive urbanization of landscapes. The latter is among the dominant anthropogenic factors influencing the natural and adaptive evolution of living populations
[2], and for this reason, it is the most important phenomenon affecting the environments in the 21st century
[3][4][5][6][3,4,5,6].
Considering the multitude of urban contexts, purposes, and needs for which green spaces in cities are created, it is today very challenging to provide an exhaustive definition of ‘urban area’ and its relative ‘urban vegetation’, since the geographic, climatic, and resource-related opportunities, and constraints, are not equally distributed factors across the world and specific for each context. Furthermore, urban vegetation can also include cultural plant typology with agricultural interest related to food production, such as the horticultural species. Starting from these considerations, the present rentryview aims to provide a general overview of how genetics and genomics notions can help in the selection and improvement of plant species and/or suitable varieties for specific urban contexts. In detail, after a brief overview of the genomic tools available and potentially usable in genetic improvement plans, researcherswe focused on the evaluation of particular genetic traits for the selection of plants in relation to the urban area in which they are used and the environmental stress to which they are generally subject. Finally, researcherswe took into consideration the contribution of genetics in achieving specific goals that are usually set in urban contexts.
Processing Criteria
With a univocal classification of urban green spaces not being present
[7][8][9][7,8,9], in order to allow a better understanding of some of the aspects covered,
reswe
archers judged it appropriate to simplify the classification of these environments by empirically dividing them into four macro areas, according to their surface availability per plant and their intended usage as follows:
- (i)
-
Urban/metropolitan parks: these are green areas in cities, and other incorporated places, that offer recreation and green spaces to residents of and visitors to the municipality;
-
- (ii)
-
Urban gardens: these are areas where urban vegetation is exploited to provide food products, especially employing horticultural species. Urban food production can be carried out by citizens or administrations in private buildings and public spaces for self-consumption, or can be performed by farms with commercial purposes, also using innovative outdoor or indoor growth systems;
-
- (iii)
-
Road verges: these are small, vegetated areas composed of grass or plants and sometimes also trees, mainly located between a roadway and a sidewalk or within roundabouts;
-
- (iv)
-
Roofs/terraces/balconies: these are small green areas located in private or public buildings. They include both surfaces partially or completely covered with vegetation (e.g., green roofs) and container gardens where plants are maintained in pots.
-
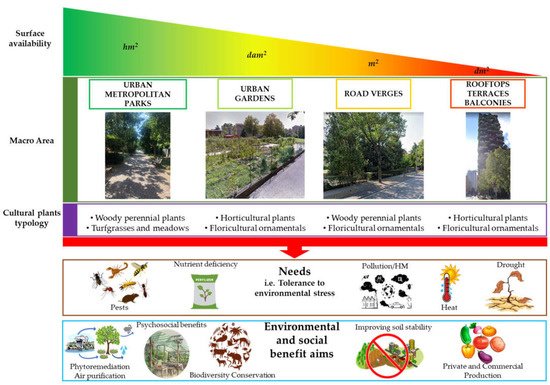
Each of these four categories is set up according to different purposes (ornamental, urban planning, bioremediation, soil consolidation purposes, etc.), that, in turn, affect the choice of the species/varieties to be used (i.e., woody perennial plants, meadows and turf grasses, horticultural and floricultural ornamental plants), and their adaptive characteristics (resistance to biotic, abiotic stress, hypogeal and epigeal habitus, minimum maintenance requirement, etc.). For this reason, as previously indicated, it is not easy if not impossible to define specific and common breeding programs for urban vegetation, since it includes both agricultural and not species with different genetic and physiological properties, and agronomical needs (Figure 1).
Figure 1.
Schematic representation of the multilayered framework of the most common contexts, here defined as Macro Areas, found in an urban environment. In each macro area, the goals to achieve are multiple and specific, depending on the characteristics and needs of every specific zone, and specific vegetation type that is desired to use. However, under all conditions, the need to use specific cultural plant typology able to cope and tolerate environmental stresses is a fundamental requirement. In addition, the potential environmental and social benefits deriving from them are indicated. Details are described in the text.
The bibliographic analysis for this work has been performed exploiting Scopus® in order to search for articles and reviews of interest written in English by integrating different combinations of selected keywords for every paragraph, including terms defining:
- (a)
-
The environment of interest: “urban areas”, “cities”, “green areas, “green gardens” “public green”, “public parks”, “urban agriculture”;
-
- (b)
-
The plant typologies: “plant”, “ornamental”, “flowering”, “horticultural”, “woody”, “trees”, “meadows”, “turfgrasses”;
-
- (c)
-
The genetic subject: “breeding” “molecular markers”, “marker-assisted selection” “marker-assisted breeding” “molecular selection” “genomic selection” “genomics” “genetic improvement” “variety” “cultivar”;
-
- (d)
-
Specific goals: “abiotic stress”, “heat shock”, “biotic stress”, “pathogen stress”, “water stress”, “drought”, “dwarf” “compacted”, “growth habit”, “edible flowers” “food production”, “leafy vegetation”, “baby leaf”, “phytoremediation”, “air purification”, “biodiversity”, “soil erosion”, “soil stability”, “psychosocial”, “ecosystem services”.
-
The lists of documents found have been then manually screened to select the most relevant works. In addition, the research has been deepened with a snowball sampling approach, exploiting citations and references included in the already screened papers.
2. Genetic Information as a Genomic Tool That Is Potentially Helpful in Breeding Approaches for Urban Contexts
Plant breeding is a science-driven creative process that involves the evaluation, selection and propagation of plant populations characterized by a combination of specific desirable traits
[10]. Originally, conventional breeding was exclusively based on time-consuming and expensive phenotyping observations: plant selection was accomplished through massive screening of phenotypic parameters mainly related to the morphology of the plant (fruits, flowers, stem), to its tolerance to both biotic (insects, molds and viruses), and abiotic stresses (temperature, drought, salinity, heavy metals)
[11].
Genetics made it possible to overcome many of the limitations associated with conventional breeding, facilitating, and speeding up the process of varietal constitution. New opportunities for crop improvement were generated thanks to the increasing availability of whole genome data that, in turn, enabled the discovery of candidate genes related to traits of interest, mutations responsible for phenotypic variability (e.g., single nucleotide polymorphisms, SNP or insertion/deletion, Indel) and molecular markers associated with traits of interest (e.g., simple sequence repeats, SSR)
[12]. Overall, these molecular data are a powerful predictive tool for the targeted selection of superior genotypes. However, as their discovery is strictly dependent on the availability of sequenced genomes, for many years, their application in breeding processes has been limited to a few sequenced species of great agricultural and economic importance, such as rice, barley, tomato, and grapevine
[13]. Moreover, before the advent of single molecule real-time (SMRT) sequencing (better known as third-generation sequencing platforms), the complex architecture of a genome represented another major obstacle to the assembly process. The set of sequenced genomes belonging to ornamental species and some tree families still only represents a small fraction of all genomes sequenced thus far for three main and often coexisting reasons
[14]. First, in this category of plants, the high level of heterozygosity along with the abundance of GC and repetitive sequences represent a significant source of ambiguity during genome reconstruction. Second, some fruit tree species are often widespread in urban areas (e.g., the
Citrus genus)
[15], and several ornamental plants (e.g.,
Rosa,
Chrysanthemum,
Lilium,
Mandevilla, and
Primula)
[16] are the result of intraspecific and interspecific crosses and ploidy manipulation. These aspects increase the probability of chimeric assemblies because of incorrect connections between scaffolds belonging to homologous chromosomes
[17]. Finally, technical problems in the assembly phase are frequently encountered in species characterized by mega-genomes (i.e., genomes > 10 Gb), such as ornamental plants (e.g., peony, 13.79 Gb)
[18] and tree genera typical of the urban context (e.g.,
Cedrus, 15–16 Gb, and
Pinus, 31 Gb)
[19].
The progressive collapse of sequencing costs and the release of tools able to overcome part of the technical challenges previously mentioned made HTS (High-Throughput Sequencing) platforms accessible also to minor crop species (also known as orphan crops, for review, see for example Simko et al.
[20] and Bohra et al.
[21]) and ornamental plants (for review, see Zheng et al.
[14]). As a matter of example, the number of sequenced genomes completed yearly for ornamental plants significantly increased from 1 in 2012 to 17-20 in the last five years, and currently, the genome sequences of more than 70 ornamental species have been released
[14]. Conversely, most of the tree species frequently found in urban green areas of the Mediterranean basin (e.g.,
Carpinus betulus,
Tilia cordata,
Acer platanoides, and
Pinus pinea) remain unsequenced today.
Genomics and transcriptomics open a wealth of opportunities not only for large-scale plant breeding but also for small-scale cultivations, such as those in an urban environment. In
Table 1 reswe
archers reported the main molecular approaches currently used to connect phenotype to genotype. In fact, the ultimate goal in plant breeding is to use the genotypic information to predict phenotypes and select improved cultivars
[22][23][22,23].
Table 1. Overview of the main genomics and transcriptomics techniques useful for breeding purposes.
|
| Techniques |
|
| General Description |
|
| To Learn More about |
|
|
| Whole Genome Sequencing (WGS) |
|
| The genome of a species is assembled for the first time into chromosomes with high coverage and it is functionally annotated to produce a reference assembly and to predict hundreds of loci underlying agronomic traits |
|
[24]
|
|
| RNA-seq analysis |
|
| RNA-seq can be used to examine the RNA sequences that are present in a sample (transcriptome). This is crucial for linking the information contained within the genome with the functional proteins that are expressed. RNA-seq can be used to elucidate which genes are turned on or off within a cell under specific conditions |
|
[25]
|
|
| Whole genome resequencing (WGR) |
|
| The genome is fully sequenced with low or modest coverage and is aligned against the reference genome assembly to predict allelic variants |
|
[26]
|
|
| Reduced |
| Representation |
| Sequencing (RRS) |
|
| GBS |
|
| A fraction of the genome is sequenced and aligned against the reference genome assembly to predict allelic variants. For GBS, ddRAD-seq, 2bRAD-seq and ezRAD-seq the regions to be sequenced are randomly chosen using restriction enzymes, for target-seq the regions to be sequenced are selected through PCR |
|
[27]
|
|
| ddRAD-seq |
|
[28]
|
|
| 2bRAD-seq |
|
[29]
|
|
| ezRAD-seq |
|
[30]
|
|
| target-seq |
|
[31]
|
Regardless of the methodology used, if the allelic variant(s) is responsible for a favorable trait, it can be subsequently introgressed into an elite variety through a combination of conventional breeding and molecular marker-assisted selection (MAS). MAS is defined as a process where molecular markers associated with a specific trait of interest are used for the selection of the trait itself. In this sense, the use of molecular markers is particularly indicated when the phenotypic evaluation of the trait is hardly doable (e.g., disease resistance) or expensive (e.g., male sterility)
[24]. MAS is also effective in selecting desirable gene alleles independently from possible confounding environmental, pleiotropic or epistatic effects, in monitoring the introgression of a desirable allele in backcrossing events, or in identifying and avoiding linkage drag effects
[32].
Considering the ratio between extragenic and genic regions, most variants identified through WGR or RRS are unlikely to underlie phenotypic effects. However, the combination of hundreds of noncoding allelic variants produces molecular profiles able to identify uniquely a given genotype. Such information is extensively used for screening populations and selecting parental genotypes in breeding programs (marker-assisted breeding, MAB), testing the purity and uniformity of commercial varieties, evaluating the stability of cultivars through generations, registering new varieties and for assessing their distinctiveness from preexisting varieties
[33].
For large-scale crop breeding (open-field and greenhouses), the marker-assisted genetic improvement techniques mentioned above are well documented
[34]. The development and incorporation of molecular markers in breeding processes could also be adopted for small-scale cultivation approaches (from urban parks to small private allotments). In the latter case, the goal would be the development of varieties with a genetic background suitable for urban needs. In this context, the contribution of genomics in identifying markers and genes responsible for plant adaptability in urban environments will play a crucial role.
3. The Role of Genetics in the Adaptability and Sustainability of Plants in Different Urban Contexts
In the development of any plant breeding program, it is necessary to define the desired variety’s ideotype, i.e., the ideal plant model endowed with every trait of interest for the specific aims and destinations
[35]. While some adaptive characteristics of plants are shared by all types of urban vegetation, there are others that must respond to specific context conditions; thus, the ideotype can change according to the macro area considered. For example, the soil requirements of a tree grown on a roadside must necessarily be distinct from those of a tree grown in an urban park. This is both to allow the plant to grow by bypassing the stress of soil deficiency and to prevent it from causing damage to the road surface as it grows. Generally, plants must be able to cope with the environment in which they are introduced, not suffering from abiotic or biotic stresses. In fact, urban areas represent environments characterized by restrictive growing conditions due to a high density of buildings, infrastructures, and heavy traffic, which cause negative effects such as soil compaction, pollution, high temperatures, drought, lack of light, presence of heavy metals, high salinity, and nutrient deficiency
[36]. The ability to cope with different kinds of stress has become even more important, considering the increasing impact that environmental changes and global warming have on plants
[37]. In past years, it has been reported that abiotic stresses reduced by as much as 50% the yields of most major crops worldwide
[37][38][39][37,38,39]. Providing varieties able to adapt to several different types of environments and resilient to stress factors is in general a major challenge in the immediate future for breeders to guarantee cultivation
[40][41][42][40,41,42]. Moreover, plants must also be resilient to stresses to limit the usage of agronomic inputs. Modern cultivars have been mainly bred for performance under a high supply of factors such as irrigation water, fertilizers, pesticides, and interventions such as tillage and manutention
[43]. Hence, the need to provide varieties environmentally and economically sustainable stands with a view of limiting costs and energy exploited for their cultivation, reducing limited resource usage, and avoiding the introduction of polluting chemical compounds into the environment
[44][45][44,45]. In addition, to guarantee plant sustainability, it is important that they do not present potentially harmful traits for ecosystems and human health, such as invasiveness and high-allergenic compound production
[46]. An important strategy to cope with the adaptability and sustainability problem is to recur to wild or traditionally used populations, diffused in areas with environmental conditions similar to those of the destination of interest. This is a key factor because, generally, these accessions are resilient to the biotic and abiotic stress typical of their diffusion area
[47][48][47,48]. In addition, not or lowly genetically improved populations present a higher genetic variability in comparison to commercial varieties, allowing to have a wider range of alleles to be drawn on for the specific breeding aims. From this point of view, the creation and maintenance of germplasm banks containing wide collections of plant material (seeds or plants conserved in vivo, in vitro or cryo-conserved) derived from wild populations and local varieties is a strategy of great relevance for having a continued availability of genotypes with useful traits
[49][50][49,50]. With regard to germplasm collections, it is important to point out that presenting a high genetic variability also means an increase in the possibility of having in the selected genotypes alleles linked to undesirable or even harmful traits, e.g., the susceptibility to a pathogen or the production of a toxic compound. These alleles can segregate with traits of interest (linkage drag), therefore, during the breeding process, careful work of selection against them is necessary
[51,52,53].To analyze more in detail the adaptability of plants to the environment, the subsequent sections provide a specific discussion about two of the most impacting abiotic stresses in urban areas as examples, high temperatures and drought, in addition to the pathogen stress.
3.1. Abiotic Stresses: Heat and Water Stress
In urban areas, the average temperature can be recorded as up to 10 °C warmer than the surrounding rural areas due to impervious surface cover, anthropogenic heat sources, and low vegetation cover. Warmer temperatures increase vapor pressure deficits, creating a greater atmospheric demand for water via transpiration and reducing soil moisture, which limits [51][52][53]the amount of water available to roots. In addition, heat stress negatively influences plant physiology as a consequence of protein damage, enzyme inactivation, photosynthesis inhibition, cell membrane deterioration, cell division interruption, and disturbance of reproductive phases [54,55,56]. Heat stress responses include the involvement of heat shock proteins (HSPs), which have a chaperone function, hence assisting the correct conformational folding of misformed proteins and helping in damaged protein movement and degradation [57]. In recent years, the molecular bases of high-temperature defense have been studied in the species of greatest horticultural interest and potentially usable in an urban context, including tomato (Solanum lycopersicum L.) [58,59], lettuce (Lactuca sativa L.) [60,61], spinach (Spinacia oleracea L.) [62,63], identifying specific transcription factors (heat stress factors—HSFs), HSPs or molecular markers related to them. Considering ornamental species, analogous results were obtained with Rododendron hainanense Merr. [64] and carnation (Dianthus caryophyllus L.) [65]. However, the use of molecular markers associated with HSPs or HSFs for breeding aims has received little attention outside of cereals. In chili pepper (Capsicum annuum L.) a study reported the introgression of two genes encoding HSP70 and HSP24 from a heat-tolerant breeding line to an elite commercial variety using markers and backcrosses with the ladder, without genetic transformation. For the tolerance-related gene selection, a MAS approach was conducted with two markers that were closely linked to the genes; while for the recovery of the recurrent parent line’s genome, to obtain the agronomic traits of interest, a MAB strategy was followed with a set of 250 paired SSRs [66].
An increase in global warming leads to enhanced evapotranspiration, reduced rainfall, and consequent water stress [67,68]. The resulting water shock for plants can also be a serious issue for urban cultivation. The solution cannot consist of more frequent irrigation due to a need in reducing water withdrawals for agriculture and in ensuring the supplies of this limited resource all over the world in the future [69]. To select plant species and to develop drought-tolerant varieties, hence requiring minor irrigation, it is important to identify the correct traits of interest, since resorting to the right phenotyping strategies can be onerous, and resistance mechanisms are plentiful and well-differentiated among species through evolution [70,71]. An important drought-resistance plant type is represented by xeromorphic succulent species, such as Cactaceae, Orchidaceae and Bromeliaceae, which endure hydric deficiency through water-storing tissues with high vacuoles (hydrenchyma), the CAM (Crassulacean Acid Metabolism) carbon fixation pathway, or waxes and cuticles on the top page of leaves [72]. These plants are often used in urban areas for ornamental purposes; hence, the introduction of breeding programs for these non-genetically improved species can be an interesting choice for breeders. For other plant types, traits of interest for drought tolerance comprehend high leaf mass per area unit, leaf thickness, unit leaf area, and leaf water potential at the turgor loss point [20,73,74,75,76]. In addition to morphological parameters, biochemical analysis of the osmoprotectant content has assumed importance if used as metabolic markers, as well as assays for the enzymes and proteins most involved in signaling drought and other abiotic pathways related to stress response, such as superoxide dismutase (SOD), ascorbate peroxidase (APX) or LEA proteins [77]. Often, morphological and physiological responses to water deficiency are in common with those for other abiotic stresses, mainly derived from heat, salinity, and coldness, but in other cases, they can be in opposition. For example, a high accumulation of proline can protect against wilting but can also lead to toxic effects in the presence of high temperatures [70,78]. Hence, attention to these possible relations must be paid if the goal is plants’ genetic improvement for multiple sources of stress. To date, the genetic bases of water deficiency tolerance have been investigated through genomic, transcriptomic, and metabolomic approaches also in species of the greatest horticultural interest but suitable in urban areas [48,79,80,81,82], and in sunflower [83,84,85]. This led to the identification of QTLs (Quantitative Trait Loci), single genes, specific sequences such as miRNAs, or genome-wide marker collections potentially related to drought traits.
3.2. Pathogen Stress
Developing plants able to resist pathogens is a very important goal for breeders since biotic stresses are among the main limiting factors for world agriculture [25,86,87,88]. In addition, there is a need to reduce the use of harmful pesticides [89,90,91,92], an important aspect, especially in urban areas where human presence is considerably high and where people are more likely to come into contact with treatment residues that pollute the environment. Compared to other traits, pathogen resistance was more often found to be controlled by a single gene than by multilocus interactions. Qualitative resistances are mostly inherited dominantly; they concern biotrophic pathogens and are effective only against particular races of pathogen species [93]. This monogenic inheritance has allowed the fixing of the trait in many varieties of species commonly found in urban areas using traditional breeding methods, but the main issue remains that the durability of the resistance is low: fungi, oomycetes, bacteria and viruses can rapidly mutate in the gene coding for the product recognized by the host, making plants return susceptible. In addition, resistant plants exert a selection pressure that favors virulent mutations in pathogens [94,95,96]. Consequently, the time required for traditional breeding is often too long to provide new resistant varieties before pests adapt to the current ones. Possible methods to overcome this problem include the use of multiline cultivars, in which every line presents different resistance genes (R genes) [97], combined with the pyramiding approach, which consists of the transfer of several R genes into the same plant [98]. MAS can be very helpful in facilitating the R gene transfer from wild or local populations, often presenting resistance as showing adaptation to the environment, to the commercial lines and hence reducing breeding times. Many diagnostic markers are available for the horticultural species of highest commercial importance, such as tomato (Solanum lycopersicum L.), and they are currently used in private breeding programs (reviewed by Foolad and Panthee [99], Lee et al. [100], Bhardwaj et al. [101], Simko et al. [20]). On the other hand, it is rare for pathogens to adapt in a short time to quantitative resistance, but it is also true that this kind of plant resistance is often difficult to introduce into commercial varieties, even by using MAS. This is because quantitative resistance has been shown to be dependent on a great number of QTLs, each with a small effect [20,97]. A modern tool that could be helpful in future breeding programs for quantitative traits and particularly for pathogen resistances is genomic selection (GS). It refers to the exploitation of high-density molecular marker collections (produced exploiting the modern NGS technologies and mapping approaches such as GBS), with statistical models to predict the breeding value for complex traits of interest [102,103].
3.3. Limited Surface Availability
The surface availability, often limited, may be the most important aspect influencing the subsequent genetic improvement programs of the chosen cultural typology. This aspect is not always transversal for all habitats since it strongly depends on the subarea considered and is specific to the different cultural typologies: for example, the surface can be high in an urban park, medium in an urban garden and/or roadside/traffic islands, and low in the roof/terrace/balcony context. In urban parks, it is possible to plant those species that have high dimensions of aerial parts and roots, such as arboreal ones; however, in roadsides, traffic islands or buildings’ terraces and balconies, the choice is more oriented toward plants able to grow in limited surfaces and, consequently, toward the selection of varieties with limited size [104,105].
To reduce the typical large sizes of trees, dwarf rootstocks were selected for the orchard species of greatest interest [106,107,108], with dwarfism being linked to reduced levels or perception of gibberellins (GA) and brassinosteroids (BR). Hence, genes related to the biosynthesis, and signaling of these phytohormones, in particular, the Wound-induced Receptor-like protein Kinase (WRK) and NAC transcription factors [109,110], are of main interest. An additional aspect that should be considered when developing varieties suitable for tight places is the growth habit. Indeed, also in the case of plants presenting dwarfism, great development of lateral shoots would require additional horizontal space, thus limiting the plant density. Studies on shoot architecture have allowed, for example, to characterize the LAZY1 gene, which is involved in the transport and signaling of auxins, promoting the upward orientation in shoots and the narrowed angles in branches [111,112], or the Tiller Angle Control 1 (TAC1) gene, which induces the outward orientation in branches [113], or genes, e.g., WEEP, leading to a weeping growth habit [111,114,115]. In addition, it is important, mainly on roadsides, to select plants in relation to the architecture of the underground parts. This is not only to facilitate nutrient uptake and to limit competition between plants, but also to avoid the risk of ruining and deforming the soil surfaces because of overgrown roots, which is a major issue in many cities’ road management [46,116,117].
Because of their generally small dimensions, the herbaceous plants for meadows and turfgrasses can be suitable for several surface availabilities, including buildings. Moreover, given the possibility of walking on them, the use of these species does not prohibit additional uses of the occupied space. The plant varieties that are the most selected for these uses generally belong to species of the Poaceae family and can be pure or derived from interspecific hybridizations [118]. Among the most common microthermal species, Lolium perenne L., Poa spp., Agrotis spp., and Festuca spp. can be found, while among macro therms, Buchloe dactiloides Nutt., Cynodon spp., Eremochloa ophiuroides Munro, and Paspalum vaginatum Sw. are present [119]. Traits of interest for turfgrasses varieties selection regard leaf fineness, rapidity in covering the surface, tolerance to trampling and mowing, slowness in sprout growing, and aesthetic aspects such as greenness and covering uniformity. Breeding programs regarding the species destinated for this use started in the 1970s and until today were carried out with continuous selection schemes, tests of progenies and intercrosses between the best-performing genotypes. For warm-season grasses, selection programs have been dominated by clonal propagation because stolons are easy for most species and interspecific progeny sterility is common [120]. Advances in genetic improvement have been relevantly focused on reducing the turf height increment rate, in finessing leaves, in wear tolerance, in environmental adaptation to different conditions and in crown-rust resistance. In contrast, the improvements that remain to be achieved are related to tolerance to other diseases, the preservation of greenness in winter and summer, and ground cover rapidity, all traits that seem to have high plasticity determined by genotype per environment (G × E) interactions. Sampoux et al. reported that the selection of plants to lessen the growth rate has probably limited the availability of the genetic resources necessary to improve the ground cover velocity and winter greenness [121].
4. What Are the Achievable Goals with the Help of Genetics?
Anthropogenic activities lead urban environments to be affected by many ecological issues of different natures, starting from pollution, which has an impact on soils as well as on water and air
[54][55][56][57][122,123,124,125]. Plants, in addition to withstanding environmental conditions and not contributing to pollution, can directly reduce the contamination already present and in general provide ecosystem services that, consequently, improve the environmental state and human health
[58][59][126,127]. The ability to offer a beneficial effect on the territory is an aspect of great interest for urban contexts that should be considered in breeding programs and urbanistic management
[55][123]. Indeed, the setting of green areas can improve slope stability reducing erosion
[60][128], it can favor faunal resettlement enhancing biodiversity
[61][129], and can reduce rainfall water surface runoff favoring infiltration into the soil
[62][130]. In addition, it has been demonstrated that plants can limit the phenomenon of urban heat islands by mitigating thermal changes, by increasing evapotranspiration, shadowing, radiation reflection and moisture traps, hence saving energy used for cooling
[63][64][131,132], keeping lower noise pollution
[65][133] and reducing the CO
2 emissions absorbing the gas through leaves
[66][134]. Another important aspect influencing the choice of a precise cultural species for specific purposes is the ability to provide plant products, particularly food, and their intended use, whether commercial or for private self-consumption. Strongly related to this last factor, surface availability is above the main determinants that can influence the possibility of having an adequate level of productivity. This is in general the largest bottleneck, as space in urban areas is often very limited and indirectly forces specific use choices and selection priorities
[67][104]. Last, although it is not a primary concern for genetic selection plans, the supply of the psychosocial benefits that are derived from the creation of green urban spots assumes an important role in cities, where the high human presence and the reduced availability of cultivated areas are typical [135].4.1. Phytoremediation
Regarding the causes of pollution in urban areas, the major responsibilities are heavy metals, such as cadmium, lead and zinc, mainly found near roads [136], and organic compounds, such as petroleum hydrocarbons (PHCs), polycyclic aromatic hydrocarbons (PAHs), chlorinated solvents, explosives and polychlorinated biphenyls (PCBs) [137,138,139]. For a long time, it has been known that several woody and herbaceous ornamental species are able to reduce the presence of these molecules in the environment, a process that is called phytoremediation, with methods including the accumulation (phytoextraction) or conversion to harmless forms (phytodegradation) of pollutants [140]. The higher dimensions allow woody plants to absorb larger quantities of pollutants by roots and from deeper soils. In contrast, herbaceous plants can be used in small spaces and can be replaced more frequently, which is a useful aspect to avoid the risk of dispersion of the absorbed pollutants [141]. Instead, more difficulties can be seen in the use of food plants for phytoremediation, unless in the case of total contaminant degradation or if the accumulation occurs in completely separate organs from those related to commercial products. Regarding woody plants, species of the genera Populus and Salix are the most interesting for phytoremediation. Nevertheless, other species, such as Ailanthus altissima (Mill) Swingie, Betula pendula Roth, Carpinus betulus L., Platanus × hispanica Mill. Ex Muenchh. “Acerifolia” and Robinia pseudoacacia [142], have been recognized as promising choices for phytoremediation purposes, although in some cases, as for Ailanthus altissima, some of their physiological properties make them invasive species. Among herbaceous ornamentals for which the capacity of pollutant presence reduction has been assessed, there are Dianthus chinensis L., Impatiens balsamina L., Portulaca grandiflora Hook., Portulaca oleracea L., Tagetes spp., and Vinca rosea L. [141]. Although many suitable species for phytoremediation in urban areas have been selected, genetic improvement programs, including phytoremediation capacity as a trait of interest, have not yet been carried out with either classical or MAB approaches. Important information on the molecular bases has been obtained in the model species not employed in urban areas Arabidopsis and rice, identifying several responsible genes of heavy metal uptake, translocation, vacuolar sequestration, and genic regulation [143]. Using nontransgenic techniques, in rice genotypes with mutations in genes related to this aspect were identified after irradiation with mutagenic agents, and varieties of this cereal were selected with MAB methods only for low accumulation of cadmium or arsenic to avoid the risk of toxic molecules’ presence in the edible products [18,144,145]. Nonetheless, these goals highlight the potential of using the same approaches to the opposite objective, hence, to obtain consistent heavy metal accumulation, and to extend this methodology also to plants suitable for urban areas. In addition to high toxic molecule uptake, accumulation, and degradation capacity, other traits of interest for future plant breeding programs for phytoremediation are size, surface and architecture of roots. They ideally would be designed to have the maximum pollutant interception capacity, given space restrictions and without incurring the risk of soil failure, especially along roadways [46]. Another important factor to consider in breeding varieties destinated to phytoremediation is avoiding the return of the absorbed pollutants to the environment. Therefore, for species that accumulate pollutants in their leaves, selecting genotypes without leaf scattering can be relevant [141].
4.2. Air Purification
The removal of CO2 and gaseous pollutants, such as carbon monoxide, ozone and nitrogen dioxide, is performed by plants through interception by their stomata, while particulate matter (PM10 and PM2.5) adheres to leaf surfaces with the aid of wind currents, where it is then absorbed [146,147]. The effects of plants in reducing atmospheric pollution in cities were studied, demonstrating their potential, particularly that of trees, in improving air conditions [148,149]. It has been reported, for both woody and herbaceous species, that plants with a high presence of leaf hair or wax cover are the most performant in capturing PMs [150,151]. Indeed, wax layers facilitate PM adhesion, while hair greatly extends the surface available to intercept composts, a factor influenced also by leaf morphology, leaf area index (LAI), and, in general, aboveground parts’ sizes. Leaf fineness and porosity are also important to favor air movement through the structure without being thrown away. In addition, the maintenance of the leaves for the entire life cycle is relevant to always guarantee pollutant removal services; hence, choosing deciduous trees can be limiting [46,152]. Finally, another aspect to consider is the single species’ susceptibility to pollutants. In fact, there are plants, such as some conifers, that are evergreen and very efficient in particulate absorption, due to their complex shoot spatial structures, but they are also particularly susceptible to these pollutants, making them possibly unsuitable for the most polluted urban areas [122,148,150]. The reported traits are very interesting in species selection and genetic improvement for air purification purposes. Among the most performative woody and shrub species, Albizia julibrissin Durazz., Betula pendula Roth., Cinnamomum camphora (L.) J. Presl, Nerium oleander L., Pinus mugo Turra, Pinus sylvestris L., Stephanandra incisa (Thunb.) Zabel, Taxus x Media Rehder, and Taxus baccata L. have been considered for their air pollution tolerance and PM absorption capacity per leaf surface unit [150,153]. As previously reported, despite their small dimensions and their low unit absorption of pollutants, herbaceous plants have anyway raison d’être, considering the high plant density for a given surface and the possibility of coupling them with trees, allowing for a synergic effect due to the absorbance of harmful molecules at different height levels and of different types [46]. As potentially interesting herbaceous species for PM removal from the atmosphere, Sisymbrium loeselii L., Polygonum aviculare L., Convolvulus arvensis L., Chenopodium album L., Achillea millefolium L., Berteroa incana (L.) DC., and Galinsoga parviflora Cav. have been reported [151].
4.3. Improving Soil Stability
Soil erosion, especially on slope surfaces, is a well-known issue in urban environments and is favored by high compaction and limited drainage capacity. This phenomenon induces hypoxia, water runoff and flooding, and increases the risk of landslides that are potentially dangerous for viability and construction [154,155]. Plants can improve soil stability mainly through the root system, which helps in several ways, such as by anchoring the ground to provide physical support, creating additional pores for oxygen and water infiltration, or supplying organic matter, which induces stable aggregate formation [156,157,158,159]. Hence, species must be selected and varieties developed researching high root dimensions, number and stiffness, as useful traits to support the soil because of hardness and tensile strength [160]. Regarding growth habits, roots that grow parallel and close to the surface improve the soil in-plane tensile strength, while roots penetrating deeper and perpendicular to the surface enhance shear strength [161]. However, it is not possible to determine the best habit regardless of soil conditions: in the case of rocky slopes with limited soil availability, plants with superficial root systems must be rewarded; in contrast, if there are adequate soil depths, deeper roots guarantee more stability, and hence, plants having them should be preferred [158]. Attention must also be paid to the canopy, which is important to avoid erosion by rainfall water. High leaf area, elevated branch presence, dense and compact growth habits allowing high plant density can lead to homogeneous soil cover, intercepting more water and gradually reaching the soil. In this context, as already reported, focusing not only on trees but also on turfgrass and shrub selection can be interesting, since different vegetation types, when combined in the same place, can provide synergic benefits, such as multiple levels of canopy to intercept water at different heights [162]. Not including Poaceae, the available literature evaluating suitable species for erosion control, such as Lantana montevidensis (Spreng.) Brinq., Lavandula lanata L., Origanum vulgare L., Rosa abyssinica R. Br. ex Lindl., and Rosmarinus officinalis L., were reviewed by Francini et al. [163].
4.4. Food Production
Plants grown with food productive aims must be subjected to different standards if their production is for commercial purposes, in the context of farms, or if they are intended for self-consumption, as in the case of public spaces and private gardens cultivations. In the first case, it is more important to develop cultivars selecting them for their yield amount and specific qualitative aspects related to products’ shelf life and subjection to food processing [164,165,166]. Many breeding plans focusing on crop productivity have been carried out during the last century by means of conventional methods. Recently, genomic information about coding regions related to production traits and associated molecular markers has become available, particularly in horticultural species [167,168,169]. This can help breeders enhance the constitution of modern varieties. Compared to productivity traits, factors such as low maintenance requirements, environmental benefits, and decorative aspects are of greater interest in horticultural species to be used in home gardens or public urban spaces than in urban commercial farms [170]. Currently, there is no evidence that horticultural cultivars destinated for self-consumption are genetically different from those employed in farms, hence there is a need in developing the former by rewarding specific traits.
As previously reported, even more in the case of commercial destinations, surface availability can be the most limiting factor in planning an urban plantation. Strategies such as vertical farming, possibly coupled with artificial lights and soilless cultivation systems, could provide solutions to obtain an adequate product quantity for farms while limiting the surface needs [171]. Instead, not having high production level requirements, the possibilities to use horticultural plants are greater when considering private and community gardens, also exploiting simple growth systems such as pots and planters. In any of the mentioned cases, it is important to adopt small-sized plants in terms of roots, shoots, or low lateral growth. For the most important horticultural species, selections for dwarf or compact phenotypes have been made. As an example, in tomato (Solanum lycopersicum L.), which is one of the most cultivated horticultural crops worldwide, bush varieties with determinate growth were selected and are some of the most common cultivar types on the market [172,173,174], even if breeding programs mainly focused on enhancing yield, abiotic and biotic tolerances, product shelf life, organoleptic properties and nutritional values [99,175,176]. A cultivar that combines small-sized mutations, SP (SELF-PRUNING), d (dwarf) and mnt (miniaturize) [177], is represented by Micro-Tom, which has attracted much attention as a model phenotype for plant studies, as it reaches 10-20 cm in height, has a short life cycle with fruits suitable for harvesting 70–90 days after sowing, and bears plant densities up to 1357 plants/m2 [178]. Even if it was originally bred for home garden purposes [178,179], the Micro-Tom variety is now commonly used in laboratory applications for dwarfism studies, as it is particularly suitable to be transformed [180] and to study its possible application for cultivation in space vehicles [181,182]. Crosses between dwarfed varieties and others characterized by high sensory qualities of the products (i.e., sweetness) can represent a possible solution to combine the organoleptic characteristics with little dimensions, as demonstrated by Scott et al. [183].
Regarding innovative indoor production systems, such as vertical farming, soilless cultivations and plant factories with artificial lighting (PFALs) [171], the breeding priorities strictly differ from outdoor systems. The controlled environments make plants less affected by pathogens and pedoclimate conditions, allowing stress resistance to be of minor importance [184]. On the contrary, high productive capacity in quantitative and qualitative terms under low light intensities or specific wavelengths can be of primary importance, being the energetic costs a big limit for artificial lighting systems [185,186]. Light-related aspects, such as photoperiod, also influence several phenological stages, such as stem elongations and flowering. Hence, developing genotypes responsive to specific manipulation of these factors can lead to advantages such as shorter productive cycles, better harvesting synchronization, and growth habits suitable for the growth system spaces [186,187]. For the obtainment of food products derived from insect-mediated fertilization, the settle of pollinators is often conducted opening the growth environments, hence enhancing the risk of pathogen introduction. For this, it can be of interest for the plant species involved, to develop apomictic cultivars, i.e., able in producing fruits without requiring fertilization [188,189].
4.4.1. Leafy Vegetables
Aiming at exploiting limited urban spaces for commercial food production or private consumption, high attention should be given to leafy vegetables, which can provide food products needing on average less surfaces and in shorter times compared to fruit species [184,190,191]. A commercial typology of great interest nowadays and for which leafy vegetables are the most popular cultures is fresh-cut production. It consists of fruits or vegetables which have undergone minimal processing after harvest (e.g., trimming, washing, decontaminating, packaging), in order to provide ready-to-eat products maintaining their fresh state. The main issue, being the processing made before passage to the distribution chain, is the higher perishability compared to non-fresh-cut products [192]. The leafy vegetables most commonly used for fresh-cut production are spinach (Spinacia oleracea L.), kale (Brassica oleracea L. var. acephala D.C.), salad rocket (Eruca sativa Mill.), wild rocket (Diplotaxis tenuifolia L.), corn salad (Valerianella locusta L.), chicory (Cichorium intybus L.), curly endive (Cichorium endivia L. var. crispum), and lettuce, especially looseleaf varieties (Lactuca sativa L. var. acephala) [193,194]. To date, lettuce is the most popular species also in vertical farming [195]. In the last decade, breeding for fresh vegetables was mostly based on improving yields and postprocessing performance in terms of shelf life, leaving other aspects to lower priority, such as biotic and abiotic resistance. Moreover, the attention was focused mainly on tolerance to high plant density and hypoxia, post-cutting recovery ability, low core length, solid midrib and good organoleptic and nutritive profiles [192,196,197]. To select cultivars with less browning susceptibility after cutting, the use of browning activity-related enzymes and volatile molecules responsible for off-odors as metabolic markers have been proposed [196]. The market increment of the ready-to-eat products allows farms to harvest leafy vegetables at a very early maturation stage, even further reducing the crop cycle, strongly enhancing the sown density and obtaining higher yields per surface unit [194].
Baby leaf lettuces such as Green L
ast, althougeaf, Red Leaf, and Lollo Red, hence cultivars particularly suitable in providing quality products in a very limited growth time, have been developed and demonstrated to be better than whole-head lettuce in terms of harvest index, simplicity of being processed, oxidation after cutting reduction due to smaller stem diameter and appreciation by consumers [198]. Developing new baby leaf varieties can be very interesting to farms but also to private urban gardens for th
eir it is not acultivation in pots and planters, especially in balconies and other tight spaces.
4.4.2. Edible Flowers
Innovative food pr
imaroduction species suitable for urban areas, whose products are having a continuously increasing commercial interest, are edible flower plants [174]. Edible flowers present innovative and original organoleptic characteristics and can be highly
nutritive for their nutrac
onceeutical values, such as the elevated content of health molecules such as antioxidants, e.g., flavonoids, phenolic acids, and alkaloids [199]. Flower
in
for genetic seleg plants are already one of the first choices for small spaces in cities due to their high aesthetic effect, but species able to provide edible flowers can combine ornamental value with the supply of innovative food products. These species include begonia (Begonia x tuberhybrida Voss), rose (Rosa spp. L.), c
hrysant
hemum (Chrysanthemum spp. L.), pansy (Viola × wittrockiana Gams), li
lac (Syringa vulgaris L.), elder (Sambucus nigra L.), and Japanese wisteria (Wisteria floribunda (Willd.) DC.) [200,201]. Breeding o
f flowerin
plansg species has focused almost uniquely on quantitative, morphological, and aesthetic aspects of flowers [45],
aside from biot
he supplic and abiotic stress tolerance. Good values for these traits were mainly reached using hybridization, also the interspecific trait, followed by vegetative propagation of the most performative individuals [202,203]. This has made the genetic background of varieties highly
hetero
f the pszygous and, in some cases, allopolyploid or aneuploid. The genomic complexity, coupled with a relatively low economic importance for single species in comparison to that of open-field crops, led to severe difficulties in molecular marker development attempts and hence in starting MAB programs [202,204]. However, in recent y
ears, the advent of NGS tech
osocial bennologies has made it possible to obtain whole-genome sequences or high-density linkage maps for the most important ornamental species, providing powerful tools for future genomic-based breeding applications [205,206,207,208]. Regarding e
dible f
its that lower plants, the biochemical characterization of products was made for the most important species [209,210,211], but there is no evidence of genetic improvement progra
ms for
e derived from t the obtainment of high organoleptic and health-related molecules content as selection criteria, which can be detected with the use of specific assays or analyzing molecular markers linked to genes controlling these aspects. The improvement of these traits in cultivars, coupled with that of products’ shelf life, with flowers being particularly delicate and their appearance preservation being significantly important, is a key factor in promoting edible flower plant diffusion in cities [200].
4.5. Biodiversity Conservation
Urban environments are known to be ch
aracterize
creation of d by low levels of biodiversity, and the increase in urbanization inevitably leads to a reduction in the variability of life in ecosystems [212]. On the contrary, the setting
of green
areas plays an important role in conserving and also improving biodiversity, favoring the resettlement of wild plants, enhancing their local genetic diversity [213], and providing the right habitat for many animal species throu
gh the supply of suitable places for
nesting and sheltering or to find food sources [214]. Improving b
iodiversity can
spots aspositively influence the ecosystem services provided in the long term, strengthening the resilience to environmental changes [215].
One of the mos
t endangered animal categories in cities withou
mes ant plants is that of pollinating insects, being nectar and pollen primary sources for their sustenance [216,217]. To set plants i
n green spaces capable of attracting insects, first, it is important
role in cities, wto focus on allogamous species with entomophilous pollination. Flower ornamental plants are the most effective, given the attractiveness they have to pollinators due to their flowers. Traits to be considered in plant breeding programs for developing varieties aimed at enhancing insect biodiversity comprehend flower size, morphology, color and scent, pollen and nectar production quantity, and flowering period longevity [129,218,219]. An important aspect on wh
ich many studie
re the hs have focused is the role of native plants in attracting insects in comparison to exotic species, but researchers have not reached a consensus on the matter [220]. The trai
ts reported can be g
h ood indicators of insect resettlement, but to assess the potential of a variety in improving pollinator biodiversity in a specific environment, it is necessary to count and recognize flowering-visiting individuals for a few years. Garbuzov and Ratnieks [129] evaluated th
e ability to attract pollinators of 32 garden flowering plant varieties, cou
man nting them over two summers in different experimental gardens in Sussex. They noted that Agastache foeniculum (Pursh) Kuntze, Lavandula x intermedia Emeric ex Loisel. var. Gros Bleau, Edelweiss and Hidcote Giant, and Nepeta x faassenii Bergmans ex Stearn var. Six Hills Giant are the most p
er
esence and the redformant plants for the aim. Although works on varietal improvement related to this aspect are not available in the literature yet, providing traits of interest and putative suitable species can favor their insertion into plant breeding plans aimed at enhancing biodiversity.
Despite the setting of green species is in general linked to biodiversity enhancement in an u
rban area, if it regards the introduc
ed availabilittion of alien plants, there is a risk of negatively influencing the ecosystem balances due to the risk of invasiveness. In fact, specific traits of the introduced varieties or species can make these plants not only problematic for environmental management as a consequence of their high spread but also predominant in the hoarding of limited resources. This confers them a major fitness advantage in comparison to native plants, reducing the survival possibilities and the reproductive success of these last [221,222]. The competition effect can influence the present biodiversity
at of cultithe interspecific level, but it may be accentuated at the intraspecific one, likely because of the more probable sharing of an ecologic niche between the introduced varieties and autochthonous populations of the same species [223]. There is a wide range of genetic traits that can make the plant inv
asive, a
ted areas are typicnd they differ depending on species and environment characteristics. However, they generally present a greater capacity for growth under specific environmental
[68]conditions (expressible with parameters, e.g., relative growth rate, leaf area index, earliness in blooming, root system development capacity), reproductive and dispersal efficiency (e.g., seed production, flowering duration, seed dispersal distance) [224,225]. These factors often match with traits of interest; hence, it may be not possible to completely avoid the risk of enhancing the invasive potential of selected plants in breeding programs. Another important aspect that can reduce biodiversity in the case of non-native plant introduction is related to intraspecific or interspecific hybridization. In fact, progeny generated through crossing between the introduced plants and native populations of the same species or of compatible species have more chances than the parental lines in presenting competitive traits and become invasive, due to higher genetic diversity, heterosis, trait fixation or trait novelty [226,227,228,229]. Therefore, if the fertilization rate within native populations is lower than that between native plants and non-native plants or hybrids, the survival of the former is threatened. Regarding ornamentals or species for which reproductive capacity is not of interest, a possible strategy to counteract this risk is to develop sterile varieties, such as exploiting odd ploidy levels, canceling their possibility of colonizing the surrounding environment [230].
4.6. Psychosocial Benefits
Finally, other important services supplied by green urban areas are the psychosocial benefits derived from their establishment. The sense of being in contact with nature due to the presence of plants in the surrounding environment indeed has several psychological effects on urban communities. For example, making people feel more relaxed and restored, enhancing social cohesion, and improving fitness through the promotion of outdoor physical activities as well as improvement of health through the reinforcement of the immunity system [135]. In general, it is very difficult to define which of the plant traits are useful to provide specific psychosocial benefits to urban communities, but it is known that visual factors such as flower color, morphology, and sizes, as well as the emission of scents, are appreciated not only for their decoration potential but also because of their contribution to the perception of higher psychological well-being. This regards most of all flowering ornamental plants, for which these traits are some of the most researched in current breeding programs [45].
5. Summary and Outlook
The first real difficulty of an urban context lies in the definition and list of diversified typologies of green spaces coexisting often in neighboring areas, created with purposes ranging from niche food production to pure ornamental appearance. For these reasons, a simplification of the system itself, with the definition in macro areas, is of primary importance to focus the attention on the various factors characterizing and influencing such environments. As mentioned in the above sections, genetic/genomic approaches have largely contributed to the development of improved varieties with resilient traits of interest, and the use of molecular markers has amply demonstrated its potential, making it one of the most useful predictive tools in genetic improvement, not only for major plants or in crop science. In fact, in the post-genomics era, there is a striking abundance of genomic resources, such as genome sequence assemblies, germplasm sequencing data and gene expression atlases, for species potentially employed as urban vegetation
[14].
This
re
ntryview is intended to evaluate the potential genetic aspects that must be considered in a breeding program for varieties or species to be grown in an urban context. These characteristics must take into account both the adaptability of the plant to the specific area where it must grow and to the purposes for which it is used. As suggested by Henderson and Salt
[69][231], studies on biodiversity, for example, through the genetic characterization of plant germplasm collections and a correct hierarchy of selectable physiological and environmental parameters, can provide a potential genetic resource from which to appeal for targeted planning. Generally, the selection and use of favorable effect alleles in breeding programs are required to enhance genetic variance and to improve the rate of genetic gain in all environmental landscapes, but even more so in an urban scenario, an integrated approach is required to realize genetic gain through the modernization of breeding programs. Thus, in such a context, the appropriate choice of parents and optimized breeding pipelines for the fixation of target alleles present potential ways to enhance breeding efficiency and developing modernized breeding programs will help realize higher genetic gains in urban contexts for developing climate changes-resilient species.
Furthermore, a research improvement in the horti-floricultural sector with new varieties to be entered into the urban market can lead to a boost in socioeconomics that encourages the spread of vegetable, flower, and ornamental species in public and private spaces, with particular reference to environments where the presence of vegetation has so far been limited. In addition, as previously described, it turns out to be an investment in terms of reduction in greenhouse gas emissions and pollution, enhancement of territorial biodiversity, protection of ecosystems, etc. More efficient development of plant varieties can also promote breeding and nursery companies, leading them to increase and diversify the range of products and initiate innovative genetic progress plans. The certification of the products obtained through registration in the official variety registers also guarantees the economic benefit, certifying the intellectual property and safeguarding against fraud.
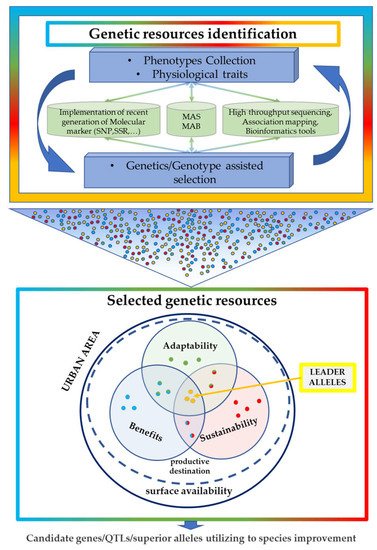
In conclusion, as summarized in Figure 2, the characterization of the urban vegetation with the best genetic profiles represents the new green challenge in urban planning to have the right genetics in the right place. A simplification and hierarchization of the main aspects that characterize an urban context, such as the surface availability, productive destination, adaptability, sustainability, and benefits contribution (details are reported in the text), play a key role in a breeding program. The identification of more appropriate starting genetic resources, by phenotype collections and physiological traits, is fundamental for the identification of appropriate traits/donors/parents and subsequent crossing programs. The potential application of molecular MAS and MAB and computational tools for performing genetic and/or genomic-assisted selection. Data generated through these trials can be used in the selection of specific leader alleles that can be introduced in species improvement programs for the constitution of new varieties that respond in a more targeted way to different needs in an urban area.
Figure 2. Graphic overview of genetic resource exploitation for plant breeding in urban plans.