Local Brazilian goat breeds comprise six main groups (Moxotó, Canindé, Repartida, Marota, Graúna, and Azul) and the non descript group derived from crosses.
Local Brazilian local goat breeds is a vital genetic resource because they represent the mainprincipal source of animal's protein to local smallholder communities in the Brazilian semiarid.
- smallholders
- local genetic resources
- Brazilian goats
- diversity
Local 1. Introduction
Many domestic species originating in Europe were introduced to America in the colonial period. Brazilian goat breeds are derived mainly from Portuguese settlers with animals since the 16th century [1][2]. The Brazilian goat breeds were developed in the national territory due to natural and artificial selection promoted by smallholders, emphasizing the morphology and fitness traits and other associated processes like genetic drift [1]. They developed unique traits such as rusticity, prolificacy, and disease resistance [3], becoming a vital economic resource to household communities.
In addition to early Iberian introductions, Asian breeds (Bhuj, Jamnapari, Mambrina, and Angorá) were introduced. Recently, goats of European breeds (Alpine, Saanen, Toggenburg, and Murciano-Granadina) and African breeds (Anglo-Nubian and Boer) have been imported into Brazil to improve milk and meat productions.
Brazil has more than ten million goat heads, according to FAOSTAT [4]. Approximately 90% of the national herd consists of animals with no defined breed pattern (NDBP) originating from indiscriminate crossbreeding between foreign and local breeds. However, there are two officially recognized local breeds (Moxotó and Canindé), and other local ecotypes (Serrana Azul, Repartida, Marota, Graúna, and Gurguéia) named according to their origin region or a particular trait. Otherwise, since 2015, these genetic resources are officially recognized as locally adapted breeds, local, or autochthonous breeds (Biodiversity Law. Law No. 13.123/15) [5]. According to a new international breed classification system, local breeds occur only in one country [4].
The local Brazilian goat breeds Moxotó, Canindé, Serrana Azul, Marota, Repartida, and Graúna, are animals of multiple functions and adapted to the climatic conditions and smallholder farming systems. They are rustic animals with less nutritional requirements, raised extensively, and survive in areas where they predominately forage foe food with low nutritional value, unsuitable for foreign breeds. These breeds are the only source of animal protein for low-income populations in the semiarid northeast. Despite the historical and social importance of these breeds, they have been undervalued for decades due to their low contribution to the national economy and their concentration in the most impoverished Brazilian region. This fact has contributed to limited scientific knowledge about these valuable genetic resources.
Another risk factor is that in the last years, the introduction of highly specialized foreign breeds in the Brazilian herds has caused a fast replacement and genetic erosion of the local ones, as observed by [6] and [7]. This situation requires conservation programs to protect the remaining genetic diversity in these local populations. Since the maintenance of these breeds is essential to guarantee an appropriate level of livestock biodiversity and the smallholder family’s maintenance.
Accurate knowledge of genetic resource diversity is fundamental for properly targeting conservation strategies and using these resources. Genetic characterization of animals through molecular markers had it shown efficient to quantify genetic variation in different populations.
2. Results and Discussion
Genetic variation within breed and fixation index (Fis) of Brazilian goat breeds are presented in Table 1. All populations showed an average allele number of over six. The mean observed heterozygosity for Brazilian breeds was higher to 0.50. These results demonstrated the high genetic diversity in the studied populations with values ranging from 0.53 (Serrana Azul) to 0.62 (Repartida). The expected average heterozygosity followed the same trend ranging from 0.58 (Serrana Azul) to 0.65 (Repartida), and the values obtained are very similar for all six breeds. The fixation index (Fis) had values under 10% except for the Moxotó breed (13%).
Table 1. The number of samples (n), the average number of alleles (Na), average observed heterozygosity (Ho), the average expected heterozygosity (He), and FIS values of the Brazilian breeds.
|
Populations |
n |
Na |
Ho |
He |
FIS (IC = 95%) |
|
Serrana Azul |
55 |
6.22 |
0.53 |
0.58 |
0.06 (−0.01–0.09) |
|
Moxotó |
60 |
6.29 |
0.55 |
0.62 |
0.11 (0.06–0.15) |
|
Marota |
68 |
6.07 |
0.58 |
0.61 |
0.04 (−0.02–0.07) |
|
Canindé |
50 |
6.15 |
0.60 |
0.64 |
0.05 (−0.01–0.08) |
|
Repartida |
52 |
6.70 |
0.62 |
0.65 |
0.07 (0.02–0.09) |
|
Graúna |
47 |
6.56 |
0.61 |
0.62 |
0.03 (−0.03–0.07) |
|
Overall |
332 |
6.32 |
0.58 |
0.62 |
|
The mean expected heterozygosity (He) of all Brazilian populations was over 0.50. The Serrana Azul was the breed with the lowest value (0.53). The matrix of DA genetic distances [8] among Brazilian breeds and FST among population pairs are showed in Table 2. The smallest genetic distance observed was between Serrana Azul and Graúna breeds, and the highest genetic distance between Serrana Azul and Canindé breeds. This situation was similar to the FST values.
Table 2. Matrix of genetic distance DA between pairs of Brazilian breeds (below diagonal) and FST between pairs of populations (above diagonal).
|
Populations |
Serrana Azul |
Moxotó |
Marota |
Canindé |
Repartida |
Graúna |
|
Serrana Azul |
|
0.084 |
0.087 |
0.105 |
0.104 |
0.036 |
|
Moxotó |
0.124 |
|
0.072 |
0.045 |
0.051 |
0.073 |
|
Marota |
0.143 |
0.110 |
|
0.097 |
0.082 |
0.066 |
|
Canindé |
0.145 |
0.100 |
0.144 |
|
0.047 |
0.089 |
|
Repartida |
0.140 |
0.086 |
0.121 |
0.097 |
|
0.085 |
|
Graúna |
0.082 |
0.130 |
0.130 |
0.144 |
0.139 |
|
The average DA among all populations was 0.122, which indicates a clear split of the different Brazilian breeds as confirmed by the average FST value (0.075). The differences can be observed in the dendrogram built through the UPGMA method (Figure 1). Two different groups were identified: the first formed by Repartida, Moxotó, Canindé, and Marota. Serrana Azul and Graúna formed the second group. The Bootstrap values ranged from 0.59 to 0.95, showing the degree of confidence in the topology.
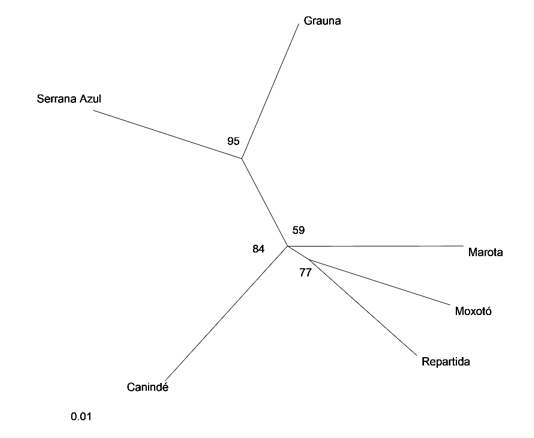
Figure 1. UPGMA dendrogram of Brazilian breeds, based on DA distance [8].
The numbers on the nodes are percentage bootstrap values from 1000 replications of resampled loci. The factorial analysis (Figure 2) showed more than 54% of the total variation in the two first axes, indicating high confidence and genetic relationship among populations. A study conducted using 13 microsatellites with six indigenous Iranian goat populations observed two main groups based on the phylogenetic tree and FCA analysis [9].
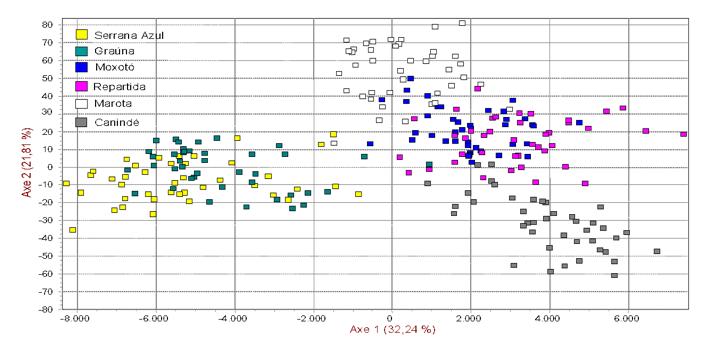
Figure 2. Spatial representation of Brazilian breeds using factorial correspondence analysis.
Ibnelbachyr et al. [10], evaluating the genetic differentiation of Moroccan goat breeds using 12 microsatellite DNA markers, found 64% of the total variation in the first three factors, whereas Bulut et al. [11] evaluating the genetic diversity of eight domestic goat populations raised in Turkey with 11 microsatellites loci, obtained 47% of the total variation of the first three factors.
The average number of alleles was higher than those found in different studies with goats in several regions globally, including some breeds in the Americas [12] and the Chinese breed of Daiyun goat [13]. However, the highest number of alleles was reported above 7 in other goat populations in Turkey [11], Marrocos [10], and Saudi Arabia [14]. The higher number of alleles presented by these studies, according to the authors, is justified by the gene flow, the location of the countries on migration routes, and the proximity of the initial centers of domestication, being the vast allelic diversity found very useful for selection. Differences in the mean number of alleles can also be influenced by the sample size and the number and type of microsatellites used.
Recent studies evaluating the genetic diversity of Brazilian goat breeds using microsatellite markers observed high levels of observed and expected heterozygosity and suggested substantial genetic diversity in the evaluated breeds.
The average heterozygosity observed in our study was higher than that reported by Silvestre et al. [15] (0.385) in Nubian goats in Brazil. On the other hand, Hossam Mahmoud et al. [14] obtained mean heterozygosity higher than 0,91, in the local Saudi Arabia goat breed.
According to Jianmin and Wenbin [16], in improved breeds, introgression and gene exchange are widespread, mainly increasing heterogeneity in the population.
Only in the Moxotó breed, the FIS value was higher than 0.10. The FIS values in the studied populations shown genetic homogeneity. The high FIS value in the Moxotó breed (0.13) may be due to structure with subdivision into herds and low gene flow among the herds. Our samples were collected from four different herds that were geographically and reproductively isolated, and each of these populations is generally closed with a reduced number of sires, which favors inbreeding. In conservation programs, inbreeding levels should be taken into account, mainly when the same male is used for a prolonged period.
The low FIS values obtained in our study were contrasting with those found by other authors in goat populations [15][17][18][19].
The degree of genetic differentiation (FST = 0.075) among all populations was predictable, recent studies have already indicated this degree of differentiation of our Brazilian goats [1], and Frankham et al. [20] suggest that FST values between 0.05 and 0.3 are typical of differentiated livestock breeds. In the European goat breeds, the FST lies around 5%.
The genetic makeup of the Brazilian goat breeds has been influenced by breeds of the Iberian Peninsula and even West African [12][21][22][23], promoting gene flow between the Iberian Peninsula, northern Africa, and the Canary Islands and Cape Verde [24][25][26]. Currently, Brazilian goat breeds are genetically distinct with a clear differentiation even from their possible ancestors [1][6][27][28][29].
As pointed out before, the Brazilian goat herds are very concentrated in one specific region, so that gene flow among breeds must be shared, especially considering that the herd book is either not ongoing or is very rudimentary. Despite this, close breeding management produces some “family” effect within breeds. In general, the average number of alleles and the mean heterozygosity obtained in this work suggest a high genetic diversity within the Brazilian goat breeds. These breeds are also targeted to crossbreeding with exotic and with each other Brazilian local goat breed, as reported by Rocha et al. [6].
The use of crosses within the local breeds is widespread in Brazil, as observed in our research (Figure 1). Those with the exotic breeds were sufficient to consider establishing a genetic conservation program for Brazilian indigenous goat populations.
Therefore, efficient herd management is vital to the conservation of these genetic resources by avoiding breeding with related individuals, exchanging individuals among herds, and increasing the effective number.
The UPGMA dendrogram (Figure 1) constructed with the DA genetic distance values grouped Serrana Azul and Graúna and performed another group joining Moxotó, Repartida, and Marota. The proximity of Moxotó and Repartida could be explained by the genetic composition of the Repartida ecotype, which arose by segregation within Moxotó, differing only in coat color.
Despite the geographical distance among the Moxotó and Canindé, these breeds probably shared the same gene pool.
The correspondence factorial analysis showed the same distribution of the populations, but in this case, the Marota and Canindé breeds were distant from the others. It was not possible to separate Moxotó and Repartida breed, and this fact was well explained before.
The Marota have had a higher genetic distance with each other breeds and, it could be due to a genetic drift effect promoted by founder effect and geographic isolation. There is a high similarity between the Serrana Azul and Grauna breeds. There was gene flow between the two groups favoring a smaller genetic distance among them. The breeders have only recently separated these populations based on the coat color pattern, and there has not been a genetic differentiation other than the few loci involved in the coat color determination.
References
- Ribeiro, M.N.; Bruno-de-Sousa, C.; Martinez-Martinez, A.; Ginja, C.; Menezes, M.P.C.; Pimenta-Filho, E.C.; Delgado, J.V.; Gama, L.T. Drift across the Atlantic: Genetic differentiation and population structure in Brazilian and Portuguese native goat breeds. Anim. Breed. Genet. 2012, 129, 79–87, doi:10.1111/j.1439-0388.2011.00942.x.
- Primo, A.T. El ganado bovino Iberico en las Americas: 500 anos despues. Zootec. 1992, 41, 421–432.
- Mariante, A.S.; Albuquerque, M.S.M.; Egito, A.A.; McManus, C. Advances in the Brazilian animal genetic resources. Genet. Resour. Inf. 1999, 25, 107–121.
- Live Animals. 2018. Available online: inhttp://www.fao.org/faostat/en/#data/QA (accessed on 3 May 2020).
- Lei Nº 13.123, DE 20 de maio de 2015. BRASILIA, DF, maio de 2017. Available online: http://www.planalto.gov.br/ccivil_03/_Ato2015-2018/2015/Lei/L13123.htm (accessed on 10 September 2020).
- Da Rocha, L.L.; Filho, E.C.P.; Filho, M.A.G.; Delgado, J.V.; Martínez, A.M.; Ribeiro, M.N. Impact of foreign goat breeds on the genetic structure of Brazilian indigenous goats and consequences to intra-breed genetic diversity. Small Rumin. Res. 2016, 134, doi:10.1016/j.smallrumres.2015.12.002.
- Ribeiro, M.N.; Da Rocha, L.L.; De Carvalho, F.F.R.; Costa, R.G.; Gomes Arandas, J.K.; Ribeiro, N.L. Threatened Goat Breeds from the Tropics: The Impact of Crossbreeding with Foreign Goats. In Sustainable Goat Production in Adverse Environments: Volume I; Springer: Cham, Switzerland, 2018; Volume 1, ISBN 9783319718552.
- Nei, M.; Tajima, F.; Tateno, Y. Accuracy of estimated phylogenetic trees from molecular data. II. Gene frequency data. J. Mol. Evol. 1983, 19, 153–170.
- Mahmoudi, B.; Panahi, B.; Mohammadi, S.A.; Daliri, M.; Babayev, M.S. Microsatellite based phylogeny and bottleneck studies of Iranian indigenous goat populations. Anim. Biotechnol. 2014, 25, 210–222, doi:10.1080/10495398.2013.850431.
- Ibnelbachyr, M.; Colli, L.; Boujenane, I.; Chikhi, A.; Nabich, A.; Piro, M. Genetic differentiation of draa indigenous breed and relationships with other goat populations assessed by microsatellite DNA markers. Iran. J. Appl. Anim. Sci. 2017, 7, 621–629.
- Bulut, Z.; Kurar, E.; Ozsensoy, Y.; Altunok, V.; Nizamlioglu, M. Genetic diversity of eight domestic goat populations raised in Turkey. BioMed Res. Int. 2016, 2016, doi:10.1155/2016/2830394.
- Ginja, C.; Gama, L.T.; Martínez, A.; Sevane, N.; Martin-Burriel, I.; Lanari, M.R.; Revidatti, M.A.; Aranguren-Méndez, J.A.; Bedotti, D.O.; Ribeiro, M.N.; et al. Genetic diversity and patterns of population structure in Creole goats from the Americas. Anim. Genet. 2017, 48, 315–329, doi:10.1111/age.12529.
- Liu, G.; Zhao, Q.; Lu, J.; Sun, F.; Han, X.; Zhao, J.; Feng, H.; Wang, K.; Liu, C. Insights into the genetic diversity of indigenous goats and their conservation priorities. Asian Australas. J. Anim. Sci. 2019, doi:10.5713/ajas.18.0737.
- Hossam Mahmoud, A.; Abul Farah, M.; Rady, A.; Mashai Alanazi, K.; Mohammed, O.; Amor, N.; Mohammed Alarjani, K. Molecular characterization of goats from Saudi Arabia using microsatellite markers. J. King Saud Univ. Sci. 2020, 32, 1681–1686, doi:10.1016/j.jksus.2020.01.001.
- De Aguiar Silvestre, E.; da Silva Costa, M.; da Silva, P.O.; Bajay, M.M.; Pinheiro, J.B.; Zucchi, M.I.; Campelo, J.E.G.; Britto, F.B. A note on the distribution of genetic diversity of Anglo-Nubian goats in central-northern farms of Piauí, Brazil. Rev. Bras. Zootec. 2015, 44, 155–160, doi:10.1590/S1806-92902015000400004.
- Jianmin, W.; Wenbin, Y. Genetic relationships of domestic sheep and goats in the lower reaches of the Yellow River based on microsatellite analysis. Biodivers. Sci. 2008, 16, 53, doi:10.3724/sp.j.1003.2008.07050.
- Hassen, H.; Rischkowsky, B.; Termanini, A.; Jessry, G.; Haile, A.; Baum, M.; Lababidi, S. Morphological and molecular genetic diversity of Syrian indigenous goat populations. Afr. J. Biotechnol. 2016, 15, 745–758, doi:10.5897/AJB2015.15062.
- Hilal, B.; Boujenane, I.; El Otmani, S.; Chentouf, M.; Piro, M. Genetic characterization of Hamra goat population in two different locations of morocco using microsatellite markers. Iran. J. Appl. Anim. Sci. 2016, 6, 901–907.
- El-Sayed, M.A.; El-Hamamsy, S.M.; Abdelhamed, W.; El-Danasoury, M.M. Genetic diversity in Egyptian goats based on microsatellite markers. Curr. Appl. Sci. Technol. 2017, 17, 130–139.
- Frankham, R.; Ballou, J.D.; Briscoe, D.A.; McInnes, K.H. Introduction to Conservation Genetics; Cambridge University Press: Cambridge, UK, 2002.
- Colli, L.; Milanesi, M.; Talenti, A.; Bertolini, F.; Chen, M.; Crisà, A.; Daly, K.G.; Del Corvo, M.; Guldbrandtsen, B.; Lenstra, J.A.; et al. Genome-wide SNP profiling of worldwide goat populations reveals strong partitioning of diversity and highlights post-domestication migration routes. Genet. Sel. Evol. 2018, 50, 58, doi:10.1186/s12711-018-0422-x.
- Silva, N.M.V.; Pimenta-Filho, E.C.; Arandas, J.K.G.; Medeiros, R.B.N.; Cavalcante-Neto, A.; Fonseca, C.; Ribeiro, M.N. Genetic relationships between local Brazilian goat breeds based on mtDNA D-loop region similarity. Span. J. Agric. Res. 2019, 17, e0407, doi:10.5424/sjar/2019174-14682.
- Amills, M.; Ramírez, O.; Tomàs, A.; Badaoui, B.; Marmi, J.; Acosta, J.; Sànchez, A.; Capote, J. Mitochondrial DNA diversity and origins of South and central American goats. Anim. Genet. 2009, 40, 315–322, doi:10.1111/j.1365-2052.2008.01837.x.
- Paim, T.D.P.; Faria, D.A.; Hay, E.H.; McManus, C.; Lanari, M.R.; Esquivel, L.C.; Cascante, M.I.; Alfaro, E.J.; Mendez, A.; Faco, O.; et al. New world goat populations are a genetically diverse reservoir for future use. Sci. Rep. 2019, 9, 1–12, doi:10.1038/s41598-019-38812-3.
- Manunza, A.; Noce, A.; Serradilla, J.M.; Goyache, F.; Martínez, A.; Capote, J.; Delgado, J.V.; Jordana, J.; Muñoz, E.; Molina, A.; et al. A genome-wide perspective about the diversity and demographic history of seven Spanish goat breeds. Genet. Sel. Evol. 2016, 48, 52, doi:10.1186/s12711-016-0229-6.
- Pereira, F.; Queirós, S.; Gusmão, L.; Nijman, I.J.; Cuppen, E.; Lenstra, J.A.; Consortium, E.; Davis, S.J.M.; Nejmeddine, F.; Amorim, A. Tracing the history of goat pastoralism: New clues from mitochondrial and y chromosome DNA in North Africa. Mol. Biol. Evol. 2009, 26, 2765–2773, doi:10.1093/molbev/msp200.
- Oliveira, J.C.V.; Ribeiro, M.N.; Rocha, L.L.; Gomes-Filho, M.A.; Delgado, J.V.; Martinez, A.M.; Menezes, M.P.C.; Bettencourt, C.M.; Gama, L.T. Genetic relationships between two homologous goat breeds from Portugal and Brazil assessed by microsatellite markers. Small Rumin. Res. 2010, 93, 79–87, doi:10.1016/j.smallrumres.2010.05.002.
- Sevane, N.; Cortés, O.; Gama, L.T.; Martínez, A.; Zaragoza, P.; Amills, M.; Bedotti, D.O.; Bruno De Sousa, C.; Cañon, J.; Dunner, S.; et al. Dissection of ancestral genetic contributions to Creole goat populations. Animal 2018, 12, 2017–2026, doi:10.1017/S1751731117003627.
- De Oliveira, J.D.; de Paiva Igarashi, M.L.S.; Machado, T.M.M.; Miretti, M.M.; Ferro, J.A.; Contel, E.P.B. Structure and genetic relationships between Brazilian naturalized and exotic purebred goat domestic goat (Capra hircus) breeds based on microsatellites. Genet. Mol. Biol. 2007, 30, 356–363, doi:10.1590/s1415-47572007000300010.